高等学校生物 > 生物I > 環境と植物
これまでの復習
編集中学や、高校生物Iの他の単元で説明した内容の復習である。 すでに読者が理解できていれば、節『植物の反応と調節』へと進んで、新たな内容を勉強すること。
導入
編集植物は一か所に固定して暮らすため、 外部環境の変化に大きな影響を受ける。 植物は外部環境の変化に対して、 自身の成長などを調節することで対応する。 このページでは、 植物と水分・光の関係、 植物の発芽・成長・花芽形成の調節、 などを扱う。
植物の生活と環境
編集環境要因
編集環境のうち生物に影響を与えるものを環境要因と呼ぶ。 植物に対する環境要因は光・水・大気(酸素・二酸化炭素)、土壌などがある。 光は、植物が光合成を行うためのエネルギー源となっている。 水は、化学反応を行う場となったり、様々な物質を輸送している。 植物は光合成だけでなく呼吸も行っている。酸素はその呼吸に必要であり、二酸化炭素は光合成に必要である。 土壌中の栄養塩類は、植物が成長するために必要である。
水分の吸収と移動
編集水は植物に必要なものの一つで、 植物は水を根の根毛から吸収し、 茎の道管を通して移動し、 葉の気孔から蒸散する。

根の外側には表皮細胞やその表皮細胞が変形した根毛があり、吸水を行っている。
その後、水は皮層の細胞やその間を通り、道管・仮道管へ到達する。
根の内部の浸透圧はその外部の浸透圧より高いので、
根は吸水する。
この圧力を根圧(root pressure)と呼ぶ。
水分子は互いに引き合う凝集力()をもっている。 この凝集力によって水は導管で途切れることなく続いている。

蒸散(transpiration)にはクチクラ蒸散と気孔蒸散があるが、ほとんどは気孔蒸散である。
蒸散量の調節は気孔の開閉によって行われる。
蒸散は、水を引き上げる力となっている。
気孔は、2つの孔辺細胞が向かい合ってできている。
孔辺細胞の細胞壁は、内側が外側より厚くなっている。
水分を吸収して膨圧が高くなると、外側に曲がり、気孔が開く。
根圧、水分子の凝集力、蒸散によって、 植物は水を吸収し移動させている。
光合成
編集植物は光エネルギーにより、 水と二酸化炭素から、 グルコースを合成している。 これを光合成(photosynthesis)と呼ぶ。
植物は光合成で二酸化炭素から酸素を作るとともに、 呼吸で酸素から二酸化炭素を作っている。 したがって、実際の光合成速度(photosynthetic rate)は、見かけの光合成速度(apparent photosunthetic rate)と呼吸速度(respiration rate)を足したものである。 イギリスのフレデリック・ブラックマンは、 光合成速度は、光の強さ、二酸化炭素濃度、温度のうち最も不足したもの(限定要因(limiting factor))によって決まるとする限定要因説()を唱えた。
光の強さと光合成速度
編集二酸化炭素濃度と温度を一定にし、光の強さを変えてみる。 光の強さと光合成速度をグラフにしたとき、 光合成速度と呼吸速度が等しく、見かけの光合成速度がゼロになる点を補償点(compensation point)と呼ぶ。 また、光の強さを上げても光合成速度がそれ以上上がらなくなる点を光飽和点(photic saturation point)と呼ぶ。
日向を好む陽生植物(sun plant)では、補償点や光飽和点は比較的高く、 弱い光でも生育できる陰生植物()では、補償点や光飽和点は比較的低い。 陽生植物にはクロマツ、ソラマメ、ススキなどがあり、 陰生植物にはブナ、コミヤマカタバミなどがある。 また、同じ植物でも、日当たりの良いところの葉(陽葉, sun leaf)は補償点や光飽和点は比較的高く、 日当たりの悪いところの葉(陰葉, shade leaf)は補償点や光飽和点は比較的低い。
二酸化炭素濃度と光合成速度
編集光の強さと温度を一定にし、二酸化炭素濃度を変えてみる。 二酸化炭素濃度と光合成速度をグラフにすると、 二酸化炭素濃度が上がるとともに光合成速度も上がるが、 二酸化炭素濃度がある一定の値以上になると光合成速度は上がらなくなる。 これは、二酸化炭素濃度が低いときは二酸化炭素濃度が限定要因となり、 二酸化炭素濃度が高いときは二酸化炭素濃度以外が限定要因となっているためである。
温度と光合成速度
編集光の強さと二酸化炭素濃度を一定にし、温度を変えてみる。 温度と光合成速度をグラフにすると、 温度がある一定の値の時に光合成速度が最も上がり、 温度が低すぎたり高すぎたりすると光合成速度は下がる。 これは、光合成を行う酵素の働きに最適な温度があるためである。
植物の反応と調節
編集発芽
編集休眠(dormancy)した植物の種子が芽を出し発育を始めることを発芽(Germination)と呼ぶ。 発芽には、水分・温度・酸素などの条件がそろうことが必要である。
休眠した種子の発芽には、水が必要である。 種子の周りには水を通しにくい種皮と呼ばれるものがあり、これが種子の休眠を維持している。
種子の休眠にはアブシシン酸(abscisic acid)という植物ホルモンが関係している。アブシシン酸は発芽を抑制する。 発芽にはジベレリン(gibberellin)と呼ばれる別の植物ホルモンが、発芽を促進している。 このようにジベレリンとアブシジン酸は、種子の発芽に関して、拮抗的(きっこうてき)に、対立する。
イネやコムギの種子では、胚がジベレリンを合成し分泌する。そしてジベレリンは胚乳の外側にある糊粉層(こふんそう)の細胞に働きかけることで、酵素のアミラーゼの発現を誘導して、アミラーゼが胚乳にふくまれるデンプンを分解することでグルコースなどの糖が生成され、これらの糖が発芽のためのエネルギー源になる。
レタス、マツヨイグサ、タバコ、シソなどは発芽に光を必要とする種子であり、光発芽種子(ひかりはつがしゅし、photoblastic seed)という。 いっぽう、カボチャ、ケイトウ、キュウリなどは発芽に光を必要としない種子であり、暗発芽種子(あんはつがしゅし、dark germinater)という。
レタスの種子(光発芽種子)は、赤色光(せきしょくこう、波長660nm)を当てると発芽を促進し、遠赤色光(えんせきしょくこう、波長730nm)を当てると発芽が打ち消される。 赤色光と遠赤色光を交互にあてた場合、最後に照射された光の波長によって発芽の有無が決まる。 最後に赤色光を当てた場合には発芽して、いっぽう最後に遠赤色光を当てた場合には発芽しない。
このような仕組みは、植物が、他に植物の多い場所では発芽しないようにするための工夫であると考えられている。なぜなら、光は植物の葉を通過すると、赤色光などは吸収されて遠赤色光だけになる。もし、他に植物が多いと、他の植物に地中の栄養や水分などを奪われやすいからである。
光発芽種子の発芽には、フィトクロム(phytochrome)という色素タンパク質が受容体として関係している。フィトクロムのように、光を受け取る受容体を光受容体(ひかりじゅようたい)という。フィトクロムには2つの型があり、赤色光を感じる型(PRまたはPrと表記)と、遠赤色光を感じる型(PFRまたはPfrと表記)がある。これらは光を吸収することによって相互に変換しあう。PRは赤色光を吸収することでPFRに変化する。PFRは遠赤色光を吸収することでPrに変化する。このフィトクロムの2つの型によって、最後に当たった光の波長が赤色光か遠赤色光かを区別している。 PFR型が増えるとジベレリンの合成が誘導され、ジベレリンによって発芽が促進される。
いっぽう、他の植物が生い茂っている場所などにある種子では、まわりの植物の葉緑体が赤色光を吸収して、吸収されなかった遠赤色光が種子に届くので、種子中のフィトクロムではPFRが遠赤色光を吸収してPRになってるため、種子中にPR型フィトクロムが多く、PFR型は少ない。
こうして種子は花芽形成や種子の発芽を調節している。
レタスの種子の発芽はジベレリンによるものなので、たとえ暗所であっても、レタスの種子にジベレリンを外部から与えれば、レタスの種子は発芽する。
成長
編集
| 刺激 | 例 | |
|---|---|---|
| 重力屈性 | 重力 | 根(+)、茎(ー) |
| 光屈性 | 光 | 茎(+)、根(ー) |
| 接触屈性 | 接触 | 巻きひげ(+) |
| 水分屈性 | 水 | 根(+) |
| 化学屈性 | 化学物質 | 花粉管(+) |
植物が刺激の方向に対して一定の方向に屈曲する性質を屈性(くっせい、tropism)と呼び、
刺激の方向に関係なく運動する性質を傾性(けいせい、nasty)と呼ぶ。
屈性には、光屈性(phototropism)、重力屈性(gravitropism)、水分屈性(hydrotropism)、化学屈性(chemotropism)、接触屈性(thigmotropism)などがある。
刺激の方向へ向かって屈曲することを正の屈曲といい、
その逆を負の屈曲という。
傾性には、傾熱性(thermonasty )、傾光性(photonasty)、傾触性(aeschynomenous)などがある。

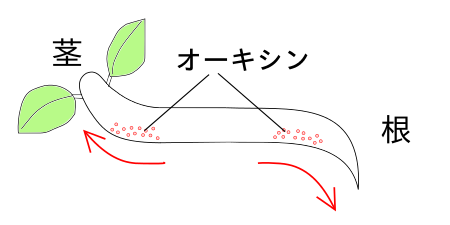
オーキシン(auxin)という植物ホルモンが、光屈性に関係している。オーキシンは茎の先端部で合成される。そして、オーキシンは光の当たらない側に移動する。そして、オーキシンの多い側(つまり光の当たらない側)では、細胞が、より伸張するため、結果的に植物が曲がる。 植物の天然のオーキシンはインドール酢酸(インドールさくさん、IAA、indole acetic acid)である。

光屈性の研究にはダーウィン、ボイセン・イェンセン、ウェント、ケーグルらの研究がある。
- ダーウィンの実験

1880年、進化論でも有名なダーウィン父子(イギリス)は、
カナリークサヨシ(学名:Phalaris canariensis)の幼葉鞘を用いて一方向から光を当てる光屈性の実験を行った。
そのまま光を当てると、光の方向に屈曲した。
幼葉鞘(ようようしょう、英: coleoptile、子葉鞘とも)を土の中へ埋め、先端部だけ土の中から出すと、先端部の下方で屈曲した。
先端部を錫箔で覆うと、屈曲しなかった。
これらから、幼葉鞘は、光の方向を感知するのは先端部であり、その刺激に反応して先端部よりも下の部位が屈曲することがわかった。
- ボイセン イェンセンの実験

1913年、デンマークのボイセン・イェンセンは、
マカラスムギ(学名:Avena sativa)の幼葉鞘を用いて一方向から光を当てる実験を行った。
先端部を切り、先端部と基部との間に、ゼラチン片を挟むと、屈曲した。ゼラチン片は水を通す。
光側に雲母片を刺すと、屈曲した。
影側に雲母片を刺すと、屈曲しなかった。
これらから、幼葉鞘は、
光を先端部で受容すると、
ゼラチン片を通る成長促進の物質が作られ、その成長促進物質は光の当たらない側に移動して、そして下方に移動して作用することがわかった。そしてゼラチン片は水を通すことから、成長を促進する物質は、水溶性であることを示唆し、実際に水溶性であることが、のちに分かっている。
- ウェントの実験

1928年、オランダのウェントは、 マカラスムギの幼葉鞘を用いて一方向から光を当てる実験を行った。 まず、先端部だけを寒天片に乗せ、一方向から光を当てる。 次に、その寒天片を光側と影側に半分に分け、 それぞれを先端部を切除した幼葉鞘に乗せる。 すると、光側の寒天片を乗せた幼葉鞘は成長しなかったが、 影側の寒天片を乗せた幼葉鞘は成長した。 これらから、先端部で作られた化学物質は、 影側へ移動してから下降し、 成長を促進することがわかった。このような植物成長の促進物質があることが分かり、オーキシン(auxin)と名づけられた。オーキシンのように、微量で植物の成長や作用を調節する物質をまとめて植物ホルモンという。 また、ウェントは屈曲の角度から成長促進物質の濃度を調べるアベナ屈曲試験法(avena curvature test)(別名:アベナテスト)を考案した。マカラスムギの学名 avena sativa(アベナ サティバ)の属名に由来する。
1934年、ドイツのケーグルは、 植物の、これらの成長促進物質をオーキシン(auxin)と名づけた。このとき、まだオーキシンの化学構造ははっきりしていなかった。
のちに植物の天然のオーキシンはインドール酢酸(インドールさくさん、IAA、indole acetic acid)という物質であることを突き止めた。
- オーキシンの極性移動

オーキシンは、茎の先端から根の方向へと移動する。逆には移動しない。これは茎をさかさまにしても、移動方向は、茎頂→根のまま変わらない。たとえば幼葉鞘を切り取ってさかさまにして、上側(根の側)をオーキシンをふくんだ寒天片に接触させても、倒立した幼葉鞘ではオーキシンは移動しない。このようにオーキシンの移動に茎→根という方向性があることを極性(きょくせい)といい、このような極性にしたがったオーキシンの移動のことを極性移動(きょくせい いどう)という。
植物の細胞膜にはオーキシンを取り入れるタンパク質(AUX1)と、オーキシンを排出するタンパク質(PIN)があることが分かっている。これらのオーキシン輸送タンパク質が、植物の器官ごとに、それぞれ細胞の特定方向の面に片寄っているので、結果的にオーキシンの極性移動が行われる。
- オーキシンと重力による移動の仕組み
-
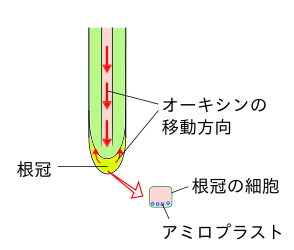 アミロプラストと重力屈性。垂直の場合。
アミロプラストと重力屈性。垂直の場合。 -
 水平の場合。
水平の場合。


オーキシンが移動する仕組みについては、まだ未解明の部分があり、学者たちの研究中である。 今のところの説は、オーキシン(インドール酢酸)は、細胞壁や細胞膜に作用していると考えられており、酸の水素イオン(H+)が関わっているとされている。
根の重力屈性の仕組みは、根冠の細胞中にあるアミロプラストというデンプンをふくむ細胞小器官が多くあり、このアミロプラストが重力によって下方に移動し、その細胞内の下部にアミロプラストが集まることが、オーキシンを輸送するオーキシン輸送タンパク質(AUX1やPIN)に、何らかの影響を与えているとされている。
- オーキシンの感受性と重力屈性
-
 オーキシンの器官ごとの感受性。
オーキシンの器官ごとの感受性。 -
 水平にした幼葉鞘の重力屈性
水平にした幼葉鞘の重力屈性


オーキシンの最適濃度は植物の器官によって異なる。 オーキシンの最適濃度は 茎>側芽>根 の順となっている。さらに、オーキシンの濃度が高すぎると、成長が抑制される。
オーキシンは極性移動とは別に、重力によって移動する。幼葉鞘を水平にするとオーキシンは重力によって下部に集まる。茎と根でオーキシンの最適濃度が違い、最適濃度を大幅に越えると、むしろ抑制されるため、結果的に茎と根が、上図『水平にした幼葉鞘の重力屈性』のように曲がって成長していく。重力と同じ方向に曲がる根のがわが正の重力屈性である。茎のがわは負の重力屈性である。
- 頂芽優勢
茎の頂芽(ちょうが、茎の先端の芽のこと)が成長しているときは、そのオーキシン濃度では側芽(そくが)は抑制されて成長できない。
これを頂芽優勢(ちょうがゆうせい、apical dominance)と呼ぶ。頂芽優勢には、サイトカイニンという別の植物ホルモンも関係している。
頂芽を除去しても切断芽にオーキシンを与えると、側芽は成長しない。また、頂芽を残しても側芽にサイトカイニンを与えると、側芽は成長する。これらの結果から仮説として、オーキシンが、側芽の成長に必要なサイトカイニンの合成を抑制している、と考えられている。
- オーキシンの光屈性
(未記述)
- その他
オーキシンの他の植物ホルモンとしては、 植物の成長を促すジベレリン(gibberellin)、 果実の成熟を促すエチレン(ethylene)、 細胞分裂を促すサイトカイニン(cytokinin)、 種子の休眠を維持するアブシシン酸(abscisic acid)などがある。
- ジベレリン(gibberellin)
ジベレリンの発見は、イネの馬鹿苗病(ばかなえびょう)という草丈の大きくなる病気の研究から、黒沢英一によって発見され(1926年)、藪田貞治郎によって単離・結晶化され命名された(1930年代)。あるカビ(学名:Gibberella、ジベレラ属)がジベレリンを分泌することが分かり、そのジベレリンがイネの草丈を大きくしていることが分かった。当初はジベレリンはカビの産生する毒素と考えられていた。その後、健康な植物自体もジベレリンを生成していることが分かり、ジベレリンは植物ホルモンだと分かった。
ジベレリンの作用は草丈を伸ばす以外にもあり、受粉してない子房に果実をつくらせ成長させる(単為結実)ので、種無しブドウなどの生産にもジベレリンは利用されている。受粉してない子房に果実作らせることを単為結実(たんいけつじつ)という。
- エチレン
エチレンは気体であり、化学式 C2H4 の植物ホルモンである。エチレンは果実の成熟をうながす。熟したリンゴからはエチレンが良く出てくる。密閉した容器に熟したリンゴと未熟なバナナを入れておくと、バナナが早く熟す。一つの箱にリンゴをいくつも入れておくと、一つでも塾すと、エチレンを出して他のリンゴも熟させるので、ほぼ同時に多くのリンゴが熟す。
気孔の開閉
編集まず、充分な水がある場合、気孔にある孔片細胞に水が取り込まれ、孔片細胞が湾曲し、結果的に気孔が開く。 水分が不足すると、葉でアブシジン酸が合成され、葉でのアブシジン酸の濃度が高まり、浸透によって後編細胞からは水が流出し、孔片細胞の膨圧が低下して気孔が閉じる。
花芽形成
編集成長すれば花となる芽を花芽(floral bud)と呼ぶ。 花芽形成には光や温度が関係している。
花芽形成が暗期の長さによって調節される性質を光周性(photoperiodism)という。
植物は一定の長さの暗期が続くと花芽形成を行い、
この一定の長さの暗期を限界暗期(critical dark period)という。
限界暗期以下で花芽を形成する植物を長日植物(long-day plant)といい、 限界暗期以上で花芽を形成する植物を短日植物(short-day plant)といい、 限界暗期に影響を受けない植物を中性植物(neutral plant)という。 長日植物にはアブラナ、ホウレンソウなどがあり、 短日植物にはダイズ、コスモスなどがあり、 中性植物にはトマト、トウモロコシなどがある。
人為的に限界暗期を短くすることを長日処理(long-day treatment)といい、 人為的に限界暗期を長くすることを短日処理(short-day treatment)という。
光周性に働きかけるホルモンは花成ホルモン(flowring hormone)と呼ばれ、フロリゲン(florigen)がある。 フロリゲンは葉で光を感知することで合成され、師管を通ることが分かっている。
花芽形成には低温にさらされることが必要な植物もある。
これを春化(vernalization)と呼び、人工的に春化することを春化処理()と呼ぶ。
春化が必要な植物には、秋まきコムギなどがある。
脚注
編集- ^ 吉里勝利ほか『スクエア 最新図説生物』第一学習社、2004年1月10日発行、p.176
参考文献
編集- 田中隆荘ほか『高等学校生物I』第一学習社、2004年2月10日発行、pp.214-243
- 『NHK高校講座 生物』第34-38回
- 生物学用語辞典 - Weblio 学問