高等学校生物 > 生物I > 生殖と発生
導入
編集生物は、生殖によって増え、発生の過程を経て個体(こたい、indvidual)となる。 生殖とは、生物の個体が新個体を作り出す働きであり、 発生とは、受精卵から成長した個体になるまでの過程である。 このページでは、 生殖の働きや仕組み、 発生の過程や仕組み、 などを扱う。
生殖細胞の形成と受精
編集有性生殖と無性生殖
編集生殖(せいしょく、reproduction)とは、生物の個体が新個体を作り出す働きである。 生殖には、親に雄(おす)と雌(めす)がある有性生殖(ゆうせい せいしょく、sexual reproduction)と、親に雄と雌がない無性生殖(むせい せいしょく、asexual reproduction)がある。
有性生殖では、親は、精子や卵のような配偶子(はいぐうし、gamete)という生殖細胞を作り、配偶子どうしが合体(接合、Bacterial conjugation)して子となる。 配偶子には、雄と雌の配偶子の形や大きさが同じな同形配偶子(isogamete)と、雄と雌の配偶子の形や大きさが異なる異形配偶子(anisogamete)がある 。 同形配偶子は緑藻類のクラミドモナスなどに見られ、異形配偶子は種子植物や動物などに見られる。 異形配偶子には、大きな卵細胞(らんさいぼう、egg cell)または卵(らん、egg, ovum)と、小さな精細胞(せいさいぼう、sperm cell)または精子(sperm, spermatozoon)がある。 卵は栄養を蓄え、精子は移動できる。 卵と精子が合体することを受精(じゅせい、fertilization)と呼び、合体したものは受精卵(じゅせいらん、fertilized egg)と呼ばれる。 有性生殖では、配偶子が遺伝的に異なるため、子は親と異なる遺伝的性質を持つ。 生殖に雄と雌が必要だが、遺伝的多様性が得られるため、環境の変化に対応できる可能性がある。
無性生殖では、親は生殖細胞を作らずに、子を増やしていく。 無性生殖には、分裂、出芽、栄養生殖、胞子生殖などがある。 分裂(fission)とは、親の体が分裂して子となる生殖の方法であり、単細胞生物のアメーバやミドリムシなどが行うほか、多細胞生物のイソギンチャクやプラナリアなども行う。 出芽(budding)とは、親の体の一部が子の体となり成長する生殖の方法であり、酵母菌やヒドラやサンゴなどが行う。 栄養生殖(vegetative reproduction)とは、植物にみられる、親の根や茎などの栄養器官(vegetative organ)が子となる生殖の方法であり、サツマイモやジャガイモやオニユリなどが行う。 胞子生殖(spore reproduction、sporulation)とは、親の体に胞子(spore)という細胞を作り、それが発芽し(germinate)て子となる生殖の方法であり、アオカビなどの菌類が行う。 無性生殖では、子は親と全く同じ遺伝的性質をもち、クローン(clone)と呼ばれる。生殖に雄と雌が出会う必要がないため効率がいいが、遺伝的多様性が得られないため、環境の変化に対応できず絶滅する可能性もある。
また、ミズクラゲのように、有性生殖と無性生殖の両方を行う生物もいる。

また、ゾウリムシは、無性生殖の分裂と、有性生殖の接合を行う。
有性生殖の目的は、環境に適応しやすくなることと、新しい核を作ることで分裂によって劣化した細胞をリセットすることである。
生殖細胞のでき方と染色体の組み合わせ
編集細胞分裂の際、細胞の核内で観察される、DNAが折りたたまれて凝縮されて棒状になったものを、染色体(chromosome)と呼ぶ。DNA(デオキシリボ核酸、deoxyribonucleic acid)とは、アデニン・チミン・シトシン・グアニン(adenine, thymine, cytosine, guanine)の4種類の塩基(base)と呼ばれるものを含む、二重らせん構造の物質である。この塩基の並び方で決定される情報を遺伝子(gene)と呼ぶ。この遺伝子の情報が、生物の形や性質を決めている。 ヒトの細胞は同形同大のペアが23組、あわせて46本の染色体をもつ。 ただし、ヒトの精子と卵は、23組のペアのうち1本ずつ23本の染色体を持っている。受精卵になると精子と卵の染色体をあわせて46本の染色体となる。
ある細胞で、ある遺伝子を決める染色体が、父に由来する染色体と母に由来する染色体の両方をもつ場合を複相(ふくそう)といい表記「2n」で表し、体細胞が例である。生殖細胞などのように、ある遺伝子の染色体が父母のどちらか片方のみに由来している場合を単相(たんそう)といい、表記「n」で表す。複相とか単相とかのことを核相という。核相は染色体の本数では決まらず、DNA量でも決まらず、ある遺伝子の染色体の種類が父母の両方由来なら2nであり父母の片方由来ならnと決まる。よって、体細胞分裂時の細胞質分裂の直前にDNA合成によってDNA量が倍化していても、核相は2nのままである。
減数分裂(meiosis)とは生殖細胞でみられる染色体数が半減する分裂である。
減数分裂は第一分裂(Meiosis I)と第二分裂(Meiosis II)の2回の分裂が連続して起こる。
減数分裂は間期(interphase)→第一分裂前期(Prophase I)→第一分裂中期(Metaphase I)→第一分裂後期(Anaphase I)→第一分裂終期(Telophase I)→第二分裂前期(Prophase II)→第二分裂中期(Metaphase II)→第二分裂後期(Anaphase II)→第二分裂終期(Telophase II)の順で起こる。



減数分裂において、DNA量の変化の時期と核相の変化の時期は異なり、一致しないので、注意。また、顕微鏡などでの見かけの染色体の本数と、DNA量にもとづく染色体の本数とが、減数分裂では一致しない。DNA量にもとづく染色体数を数えるとき、染色分体(せんしょくぶんたい)という。
- ※ このような染色体の数え方のややこしさのため、学習者は、DNA量と核相との関係について注目して、減数分裂の過程を追っていくのが簡単で良いだろう。
- 間期 染色体が複製される。よってDNA量は2倍になる。核相は2nのままである。なぜなら染色体の本数が複製で増えても、染色体の種類の数は何も変わらないので、間期が終わっても核相は2nのままである。
- 第一分裂前期 染色体が凝縮し、相同染色体が対合(たいごう)し、二価染色体(にかせんしょくたい、bivalent chromosome)となる。核相は2nのままである。二価染色体の説明で「4n個の染色分体」ともいうが、核相は2nのままであるので混同しないように。
- 第一分裂中期 前期~中期に紡錘体が形成され、中期には二価染色体が赤道面(せきどうめん、equatrial plane)に並ぶ。核相は2nのままである。
- 第一分裂後期 二価染色体が対合面で分離し、分離して出来た相同染色体は両極へと移動する。このように核分裂し、それぞれの核の染色体の数は母細胞の半分になる。DNA量は生殖母細胞と同じ。核相は n になる。
- 第一分裂終期 染色体が崩れ、核膜が形成され、細胞質分裂が起こる。この結果、核相 n の細胞が2個できる。
- 第二分裂 体細胞分裂と、ほぼ同様に分裂する。染色体の数は第一分裂の終期のまま複製されないので、最終的にDNA量は生殖母細胞の半分になる。
核相は、第二分裂の間は前期から終期まで 核相=n のままである。
- 第二分裂前期 染色体の数は第一分裂の終期のまま複製されない。
- 第二分裂中期 核膜は消失し、紡錘体・紡錘糸が形成され、染色体が赤道面に並ぶ。
- 第二分裂後期 染色分体が両極へと移動する。
- 第二分裂終期 染色体が両極に到達して、紡錘糸は消失し、染色体が糸状になり、核膜ができ、細胞質分裂が起きる。よって、最終的に4個の生殖細胞ができる。それぞれの細胞のDNA量は生殖母細胞の半分である。核相はnである。

減数分裂で二価染色体ができているとき、ある確率で、4本の染色分体のうちの、相同染色体の2本が組み換わることがある。これを組換え(くみかえ)または乗換え(のりかえ)という。(※ 生物IIで、組換えについて詳しく扱う。)
DNAの塩基配列が少し異なっていて、異なった遺伝子の情報となることがある。この違いが、個人の違いとなる。 また、性の決定に関与する染色体を性染色体(sex-chromosome)と呼ぶ。 ヒトの男女の違いは、X染色体とY染色体を1つずつ持っていれば男性で、X染色体を2本持っていれば女性となる。生殖細胞の精子は22本+X染色体または22本+Y染色体の場合があり、卵は22本+X染色体の場合だけである。つまり、X染色体をもつ精子とX染色体をもつ卵が受精すれば女性に、Y染色体をもつ精子とX染色体をもつ卵が受精すれば男性になる。
ヒトの場合、男性ホルモンや女性ホルモンなど性ホルモンを異性の人体に投与しても、ヒトの性別は変わらない。ヒトの性を決定するのは、遺伝子の性染色体である。 (※ ホルモンについて、詳しくは、単元環境と動物の反応などで扱う。)
また、ヒトの生殖細胞は、46本の染色体のうち、23組のペアから1本ずつ受け継ぐため、その組み合わせは、2^23=8388608(約800万)通りとなる。さらにその精子と卵の組み合わせは、8388608*8388608=70368744177664(約70兆)通りとなる。兄弟姉妹の違いが生まれるのは、この染色体の組み合わせの多さによる。また、組み換え(Recombination)とよばれる染色体の部分的交換により、その組み合わせはさらに増える。
動物の生殖
編集動物の生殖では、水中の動物の多くは、母体外で受精を行い(体外受精)、陸上の動物の多くは、交尾により母体内で受精を行う(体内受精)。この節ではヒトの生殖細胞を中心に扱う。 発生(development)の初期に存在している生殖細胞のもとになる細胞を始原生殖細胞(しげんせいしょくさいぼう、primordial germ cell)と呼ぶ。始原生殖細胞の核相は2nである。

1.一次卵母細胞、2.二次卵母細胞、3.精細胞、4.精子

男性では、始原生殖細胞が受精後3週目に出現し、その後体細胞分裂で増殖し精原細胞(せいげんさいぼう、spermatogonium)となる。精原細胞の核相は2nである。青年期以降、体細胞分裂を繰り返して増殖し精原細胞から一次精母細胞(primary spermatocyte)となり、減数分裂の第一分裂で2個の二次精母細胞(secondary spermatocyte)となり、減数分裂の第二分裂で4個の精細胞となる。精細胞の核相はnである。その後、精細胞が変形して精子となる。
精子は、頭部(head)、中片(mid piece)、尾部(tail)で構成される。頭部は、核(nucleus)とそれを覆う先体(せんたい、acrosome)からなる。中片はミトコンドリア(mitochondria)と中心粒(centriole)からなる。尾部はべん毛(flagellum)からなる。
ミトコンドリアは、ATPの反応によって鞭毛をうごかすことで、精子を動かすためのエネルギー源を供給する。
頭部にある先体はゴルジ体が変形・由来したものであるので、よって精子に通常のゴルジ体は含まれていない(※ 2015年センター試験の選択肢で、精子にゴルジ体が含まれていないことまで知識を問う出題あり)。

女性では、始原生殖細胞が受精後5週目に出現し、その後体細胞分裂で増殖し卵原細胞(oogonium)になる。やがて卵原細胞の多くは、退化・消失し、出生時には約200万個の一次卵母細胞(primary oocyte)ができ、青年期には約40万個に減少する。排卵の直前に、減数分裂の第一分裂で、一次卵母細胞は大型の二次卵母細胞(secondary oocyte)と小型の第一極体(first polar body)に分裂する。減数分裂の第二分裂で、二次卵母細胞は卵と第二極体(second polar body)に分裂する。やがて極体は退化して消滅する。

精子と卵が合体することを受精と呼び、生じた卵を受精卵と呼ぶ。受精の前。

ウニ(urchin)の卵を例に説明する。卵の表面には細胞膜があり、さらに外側には細胞膜ごと卵をつつむようにゼリー状の透明な層があり、ゼリー層(jelly coat)という。ゼリー層の下に卵黄膜(らんおうまく、vitelline membrane)がある。精子がゼリー層に到着すると、精子の先体に変化が起きる、これを先体反応(せんたいはんのう、acrosome reaction)という。まず先体から加水分解酵素が放出されゼリー層を溶かす。精子頭部の先体が変形し、先体から糸状の突起が出る。この突起のことを、先体突起(せんたいとっき)といい、アクチンフィラメントの束が先体突起の中身である。この働きは、ゼリー層にふくまれる物質の働きによる。このような、先体の一連の反応を先体反応という。そして精子はゼリー層を貫通する。先体突起の表面にはバインディンというタンパク質があり、ウニ卵の細胞膜にはバインディンと結合する受容体がある。バインディンと受容体が結合して、精細胞と卵細胞が融合し、受精する。1つの精子が卵に受精したとき、卵のカルシウムイオン濃度が上昇し、このイオン濃度変化によって卵の細胞膜下にある表層粒が内容物を細胞膜と卵黄膜の間に放出する。これを表層反応という。この表層反応によって、卵黄膜が硬化し、精子の侵入点を中心に卵の表面の性質が変化していき、卵黄膜が変化し受精膜(fertilization membrane)となって、受精膜が持ち上がり、受精膜が卵の表面全体に広がり、これによって他の精子の侵入を妨げる。また、卵は受精膜で保護される。このため、精子は、1個の卵にふつう1個しか受精しない(多精拒否、たせいきょひ)。受精して卵の中に入った精子は、頭部が精核(sperm nucleus)となって卵核(egg nucleus)と合体して、核相は2nとなり、受精が完了する。
被子植物の生殖
編集
植物(plant)の生殖では、カテンソウなどは風が媒介して受粉(pollination)する花であり(風媒花)、ツユクサなどは雄ずい(ゆうずい)の花粉(pollen)が同株の花の雌ずい(しずい)に受粉する(自家受粉)。この節では被子植物の生殖細胞を中心に扱う。
花は植物の生殖器官である。花には、中央に雌ずい(しずい、pistil)があり、その周りに雄ずい(ゆうずい、stamen)がある。雌ずいの膨らんでいるところは子房(しぼう、ovary)と呼ばれ、その中のつぶつぶを胚珠(はいしゅ、ovule)と呼ぶ。雄ずいの先端には花粉(かふん、pollen)を含むやく(anther)がある。

胚珠の中に卵細胞と呼ばれる細胞が、花粉の中に精細胞(sperm cell)と呼ばれる細胞がそれぞれ形成される。
花がつぼみの頃、やくの中で、花粉母細胞(pollen-mother cell)(核相:2n)と呼ばれる細胞が、減数分裂を行い、花粉四分子(かふん しぶんし)という4個の細胞になる。花が咲く頃、花粉四分子は、体細胞分裂を行い、大きな花粉管細胞()(核相:n)と小さな雄原細胞(generative cell)(核相:n)からなる花粉になる。雌ずいの先に雄ずいの花粉が付くことを 受粉(じゅふん、pollination) と呼ぶ。受粉すると、花粉から 花粉管(かふんかん、pollen tube) と呼ばれる管が伸びて、胚珠の 珠孔(しゅこう、micropyle) に到達する。雄原細胞が花粉管の中を移動し、分裂して2個の精細胞(n)と呼ばれる細胞になる。
胚珠(はいしゅ)では、胚のう母細胞(はいのうぼさいぼう、embryo-sac mother cell)(核相:2n)と呼ばれる細胞が、減数分裂を行い、生じた4個の細胞のうち、1個が胚のう細胞(embryo-sac cell)(n)と呼ばれる細胞になり、残りの3個の細胞は退化して消滅する。胚のう細胞は、3回の核分裂を行った後、細胞質分裂を行って、7個の細胞と8個の核からなる胚のう(はいのう、embryo sac)になる。胚のうは、珠孔側に1個の卵細胞(らんさいぼう)(n)と2個の助細胞(じょさいぼう、synergid)(n)、反対側に3個の反足細胞(はんそくさいぼう、antipodal cell)(n)、中央に2個の極核(きょくかく、polar nucleus)(n)を含む中央細胞(ちゅうおうさいぼう、central cell)から構成される。
- 重複受精(じゅうふく じゅせい)
被子植物の場合の仕組みである。まず、花粉管内では雄原細胞が分裂して2個の精細胞(n)となっている。そして、胚のうに達した2個の精細胞(n)のうち、1個の精細胞は卵細胞と受精し受精卵(2n)になり、もう1個の精細胞は中央細胞の極核の2個(n+n)と受精して胚乳核(3n)になる。2つの受精が起こるのでこれを重複受精(じゅうふくじゅせい、double fertilization)と呼び、被子植物のみに見られる仕組みである。
重複受精(被子植物)
- 卵細胞(n) + 精細胞(n) → 受精卵(2n)
- 中央細胞(n+n) + 精細胞(n) → 胚乳(3n)
- 花粉管の誘引 (参考)
受粉時の花粉管の 胚のう への誘引は、胚のうにある助細胞が花粉管を誘引する物質を出していることが、日本の東山哲也らの研究(レーザーで助細胞を破壊するなどの実験)によって分かっている。トレニアという植物で実験された。トレニアでは胚のうが珠皮から出ているので観察しやすいためである。
1つの 胚のう では、助細胞は2個ある。
実験結果では、助細胞を2個とも破壊すると、花粉管が、まったく誘引されなくなる。助細胞以外の、卵細胞や極核などを破壊しても、花粉管は誘引される。助細胞を1個だけ破壊すると、花粉管の誘引の確率が下がる。
そして、花粉管を誘引している物質は、あるタンパク質であることが分かっており、ルアーと名づけられた。魚釣りの疑似餌(ぎじえ)の「ルアー」が名前の由来である。このタンパク質が、助細胞で発現している。
- 裸子植物の受精

精細胞の形成:2.やく、5.花粉母細胞、7.花粉四分子、9.花粉
卵の形成:3.胚珠、4.子房、6.背のう母細胞、8.背のう細胞と極体、10.背のう
受精後:11.種子、12.胚乳、13.成熟した種子
発生とその仕組み
編集発生と分化
編集受精卵(embryo)から成長した個体になるまでの過程を発生(embryogenesis)と呼ぶ。例えば、ニワトリの雌は1日に1個程度の卵を産む。交尾をしないでも卵は産まれるが、孵化(ふか)しない。交尾をしないで受精しないで産まれた卵を無精卵と呼び、交尾をして受精して産まれた卵を有精卵と呼ぶ。無精卵と有精卵をニワトリの体温と同じ37℃で保温すると、無精卵は変化しないが、有精卵は2日程度で血管が3日程度で心臓が形成され、7日程度で脳や目や手足などが形成され、20日程度で生まれヒヨコになる。血管や心臓が発生の初期に形成されるのは、卵黄(らんおう、yolk)にある栄養を血管や心臓で取り入れるためである。
受精卵は体細胞分裂を繰り返して成長するため、それぞれの細胞は受精卵の遺伝子を全てそのまま受け継ぐ。発生の過程で、それぞれの細胞は遺伝子の異なる部分を使うことで、それぞれ異なる細胞になっていき、これを分化(differentiation)と呼ぶ。つまり、個体の全ての細胞は同じ遺伝子をもつが、使う遺伝子の組み合わせで異なる細胞になっていく。
動物の発生
編集卵の種類と卵割の種類
編集
受精卵は体細胞分裂を繰り返して成長するが、その体細胞分裂を卵割(らんかつ、cleavage)と呼ぶ。 卵割で生じた細胞を割球(かっきゅう、blastomere)と呼ぶ。
卵の極体を生じた側を動物極(どうぶつきょく、animal pole)と呼び、 その反対側を植物極(しょくぶつきょく、vegetal pole)と呼ぶ。 卵は栄養のある卵黄(らんおう、yolk)を含み、 卵黄は卵の種類によって量や分布が異なっており、 卵はその量や分布により等黄卵(とうおうらん,isolecithal egg)、端黄卵(たんおうらん、telolecithal egg)、心黄卵(しんおうらん,centrolecithal egg)に分けられる。 等黄卵(isolecithal egg)は、卵黄が少なく卵内にほぼ均一に分布しており、ウニや哺乳類などが等黄卵である。 端黄卵(telolecithal egg)は、卵黄が植物極に偏って分布しており、両生類などが端黄卵である。
卵割には、卵全体が分裂する全割(holoblastic cleavage)と、卵の一部分が分裂する部分割(meroblastic cleavage)がある。 全割には割球の大きさがほぼ等しい等割(equal cleavage)と割球の大きさが等しくない不等割(unequal cleavage)があり、 部分割には動物極側にある胚盤の部分だけで行われる盤割(discodial cleavage)と表面の細胞層だけで行われる表割(superficial cleavage)がある。
ウニの発生と分化
編集



ウニの発生は、受精卵→2細胞期→4細胞期→8細胞期→16細胞期→桑実胚期→胞胚期→原腸胚期→プリズム形幼生→プルテウス幼生→成体の順で起こる。
- ~8細胞期 等割を行う。
- 16細胞期~ 不等割を行う。
- 桑実胚期 割球が桑の実のように塊状になった胚を桑実胚(そうじつはい、morula)と呼ぶ。胚の内部に卵割腔(らんかつこう)という空所が生じる。
- 胞胚期 卵割が終わってから原腸形成が開始されるまでの時期の胚を胞胚(ほうはい、blastula)と呼ぶ。卵割腔は大きくなり胞胚腔(blastocoel)になる。胚の表面に繊毛(せんもう)を生じる。
- 原腸胚期 胞胚の次の胚を原腸胚(げんちょう、gastrula)と呼ぶ。胚の植物極側の原口(げんこう、blastopore)と呼ばれる部分から、細胞が内側に向かってくぼみはじめ、これを陥入(invagination)と呼ぶ。原口は、将来は肛門になる。原口の将来は、口ではないので注意。陥入によって胚の内部に原腸(げんちょう、archenteron)と呼ばれるくぼみができる。胚は、外表面の外胚葉(がいはいよう、ectoderm)、内側の内胚葉(endoderm)、その中間の中胚葉(mesoderm)から構成される。
- プリズム形幼生 プリズム形をしているためプリズム形幼生と呼ばれる。原腸が消化管へと分化し、原腸の先端から口もできる。発生での校門と口の順序は、まず肛門が先にできてから、あとから口ができることになる。
- プルテウス幼生 腕が生えて増えていく。
- 成体 変態してウニになる。口を下に、肛門を上にして生活している。
原腸胚のころになると、胚葉は、外胚葉、中胚葉、内胚葉に分化する。 その後、外胚葉は表皮や神経などになり、中胚葉は筋肉や骨片などになり、内胚葉は腸などになる。 [2]
- 参考画像(ウニに限らず)
-
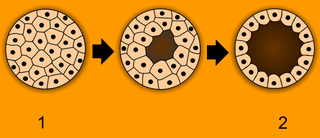 1.桑実胚、2.胞胚
1.桑実胚、2.胞胚 -
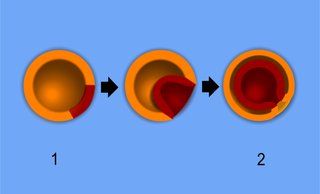 胚の原腸形成の仕組み
胚の原腸形成の仕組み
1.胞胚、2.原腸胚


カエルの発生と分化
編集
カエル(frog)の受精では、精子は動物極側から侵入する。精子が卵に侵入した位置の反対側には、灰色の部分が三日月になっている箇所が生じる。これを灰色三日月(はいいろ みかづき)という。発生が進むと灰色三日月の位置に原口(げんこう)が生じる。
カエルの卵は、卵黄が植物極側に片寄った端黄卵である。 カエルの発生は、受精卵→2細胞期→4細胞期→8細胞期→16細胞期→桑実胚期→胞胚期→原腸胚期→神経胚期→尾芽胚→おたまじゃくし→成体の順で起こる。
- ~4細胞期 等割を行う。
- 8細胞期~ 不等割を行う。動物極の割球が小さい。
- 桑実胚期 動物極側に卵割腔という空所が生じる。
- 胞胚期 胞胚腔(ほうはいこう)が、動物極側に偏った位置にできる。
- 原腸胚期 灰色三日月のあった所に、半月上の溝ができ、原口となる。この原口が陥入して、動物極の方に陥入し、原腸ができる。発生が進むに連れて原腸が拡大する。原口の上側の位置を原口背唇(げんこうはいしん)といい、主にこの部分が陥入していく。外胚葉・内胚葉・中胚葉が、それぞれできる。原口は陥入が進むにつれて弓形から円形へと変わり、表面から見ると円形の卵黄栓(らんおうせん)ができる。
- 神経胚期 外胚葉の背側に生じる肥厚を神経板(しんけいばん、neural plate)と呼ぶ。神経板はやがて管状になり、これを神経管(しんけいかん、neural tube)と呼ぶ。神経管は将来、脳や脊髄になる。神経板ができてから神経管ができるまでの胚を神経胚(しんけいはい、neurula)と呼ぶ。
- 尾芽胚 尾ができはじめた胚を尾芽胚(びがはい、tail bud)と呼ぶ。
- おたまじゃくし(幼生) 独立し食物をとる。
原腸胚のころになると、胚葉は、外胚葉、中胚葉、内胚葉に分化する。
神経胚のころになると、外肺葉は表面を覆う表皮(epidermis)と管状体の神経管に分化し、中胚葉は支持器官の脊索(notochord)と体節(somite)と腎節(nephrotome)と側板(abdominal plate)に分化し、内胚葉は管状の腸管(enteron)に分化する。
その後、外胚葉性の器官では、表皮は皮膚の表皮、眼の水晶体や角膜、口や鼻の上皮に分化し、神経管は脳や脊髄、眼の眼胞や網膜に分化する。
中胚葉性の器官では、脊索は退化し、体節は脊椎骨・骨格・骨格筋、皮膚の真皮に分化し、腎節は腎臓や輸尿管に分化し、側板は心臓などの内臓、血管の結合組織や筋組織に分化する。 内胚葉性の器官では、腸管は前部が気管・肺、食道、胃、肝臓、膵臓に分化し、中・後部が小腸、大腸、膀胱に分化する。
植物の発生
編集
胚珠内で、受精卵は発生をはじめ、珠孔と反対側の細胞は胚球()や胚柄()となり、珠孔側の細胞は吸器細胞()となる。 胚球は子葉(cotyledon)・幼芽(plumule)・胚軸(hypocotyl)・幼根(radicle)からなる胚(embryo)となり、胚柄は退化する。 中央細胞は養分を蓄えた胚乳(endosperm)となる。胚乳の養分はデンプンなどである。 珠皮は種皮(seed coat)となる。 助細胞や反足細胞は退化する。 胚珠は種子(seed)と呼ばれるようになる。
種子には有胚乳種子(ゆうはいにゅうしゅし)と無胚乳種子(むはいにゅうしゅし)がある。 有胚乳種子(Albuminous seed)にはイネやムギ・トウモロコシがあり、胚乳が発達し、発芽に必要な養分が胚乳に蓄えられる種子で、カキ科やイネ科の植物の種子が有胚乳種子である。 無胚乳種子(exalbuminous seed)にはナズナやマメやクリがあり、種子の成熟時に胚乳の養分を子葉が吸収するため胚乳は発達せず、養分が子葉に蓄えられる種子で、マメ科やアブラナ科の植物の種子が無胚乳種子である。
種子が芽を出すことを発芽(はつが、Germination)と呼ぶ。適切な水分・温度・空気などが、そろうと、発芽する。 発芽した種子では、有胚乳種子は胚乳の養分を、無胚乳種子は子葉の養分を、というように蓄えた栄養を使って成長する。 やがて葉ができると、自分で光合成して栄養を作るようになる。
コケ植物・シダ植物の生殖
編集
コケ植物・シダ植物で、胞子生殖は無性生殖である。 コケ植物・シダ植物では、胞子体(ほうしたい)をつくって無性生殖をする世代と、配偶体(はいぐうたい)という卵と精子をつくって有性生殖をする世代とを、交互に繰り返す。 このような異なる生殖方法の交代の繰り返しのことを世代交代(せだい こうたい)という。世代交代の様子を図などで環状に表したものを生活環(せいかつかん)という。
普通、胞子体は核相が2nであり、配偶体の核相はnである。なので、世代交代での胞子体と配偶体との交代にともなって、核相も交代することになり、このような核相の交代を核相交代(かくそう こうたい)という。
スギゴケなどのコケ植物で、通常に目にする植物体は、配偶体(核相n)である。コケ植物の配偶体には雄と雌との区別があり、それぞれ雄株(おかぶ)あるいは雌株(めかぶ)という。
胞子が成長して雄株または雌株になるわけだから、つまり胞子には雄雌の区別があり、雄株になる胞子と、雌株になる胞子との区別がある。
胞子をつくる胞子体の胞子嚢(ほうしのう)の中で減数分裂をして、胞子(核相:n)がつくられる。
発生の仕組み
編集発生の仕組み
編集17~18世紀頃には、精子や卵の中に、成体を縮小した形態(ホムンクルス, homunculus)があり、それが発生とともに展開するという考えである前成説(preformation theory)が有力な学説であった。それに対して、精子や卵の中に、成体を縮小した形態は含まれておらず、発生の過程で、しだいに単純な状態から複雑な状態へと成体の構造が生じてくるという考えを後成説(epigenesis)と呼ぶ。ドイツのカスパル・ヴォルフのニワトリの発生の研究などにより後成説の正しさが次第に認められていった。

割球を分離しても完全な胚になる卵を調節卵(regulation egg)と呼び、2細胞期のウニ・イモリ・カエルなどの卵が調節卵である。それに対して、割球を(ヒモで強く縛る等して)分離すると不完全な胚になる卵をモザイク卵(mosaic egg)と呼び、クシクラゲなどの卵がモザイク卵である。ただし調節卵であっても、ある程度発生が進むとモザイク卵となる。つまり、調節卵とモザイク卵の違いは、卵の各部分の発生運命がいつ決まるかの違いである。
実験には、ドイツのウィルヘルム・ルーのカエルを用いた実験、ドイツのハンス・ドリーシュのウニを用いた実験、ドイツのハンス・シュペーマンのイモリを用いた実験などがある。(後述)
なお、モザイク卵を得るために割球を縛る実験では、割球は強く縛らなければならない。縛り方が弱いと、実験は失敗する。(※ 2014年の生物Iの本試験で出題)
- シュペーマンの実験 - イモリ胚の分割実験(1902年)

イモリ胚をきつくしばる分割実験では、実験結果から灰色三日月をふくんだ部分のみが正常な幼生になることが分かった。次のような実験結果になった。
- 灰色三日月を二等分するように強くしばると、2個の胚とも正常に発生する。
- いっぽう、灰色三日月が片側の胚のみにあるように強くしばると、灰色三日月がある側の胚しか発生しない。
このことから、灰色三日月は、正常な幼生になるのに必要な物質をふくんでいることが分かる。イモリ胚の分割実験では、強くしばった場合、2個の個体になる。 なお、弱くしばると、頭が2つある1個の個体になる。この灰色三日月の部位には、背を発生させるのに必要な因子があることが、他の実験から分かっている。
予定運命
編集- フォークトの実験 局所生体染色法



ドイツのウォルター・フォークトは1925年ごろ、イモリの胚を無害な色素(ナイル青や中性赤など)を含んだ寒天片で染め分ける局所生体染色(localized vital staining)と呼ばれる手法を用いて、胚の表面の各部分が、将来どの器官に分化するかを調べた。そして、実験結果から、表面の各部がどう分化するかをまとめた原基分布図 (予定運命図) を作った。
-
 イモリ胞胚の原基分布図。側面図。
イモリ胞胚の原基分布図。側面図。 -
 イモリ胞胚の原基分布図 背面図
イモリ胞胚の原基分布図 背面図


これによると、胚の時期から、胚のどの部分が成体のどの器官に将来、分化するか決まっている。
原基(げんき、anlage)とは、まだ分化していない状態の細胞群のうち、発生段階で将来ある器官になることに予定されているもののことである。
- シュペーマンの実験 - イモリ胚の交換移植実験(1921年)

シュペーマンは、スジイモリとクシイモリの初期原腸胚で、予定神経域と予定原腸域とを交換移植してどうなるかを実験した。実験結果は、移植先の予定運命にしたがって分化した。
- 予定表皮域の移植片を予定神経域に移植した場合。 → 移植片は神経板になる。
- 予定神経域の移植片を予定表皮域に移植した場合。 → 移植片は表皮になる。
しかし、神経胚のときに移植した場合は、結果が違った。移植片それぞれの予定運命どおりに分化した。
- 予定表皮域の移植片を予定神経域に移植した場合。 → 移植片は表皮になる。
- 予定神経域の移植片を予定表皮域に移植した場合。 → 移植片は脳または神経になる。

このことから、イモリで予定運命の決定をする時期は、原腸胚初期よりは後で、神経胚になるまでには決定していることが分かる。 シュペーマンはさらに後期原腸胚でも同様の実験を行った。その結果、移植片は移植先の予定運命には従って変更される場合と、従わなずに変更されない場合とがあった。移植片の予定運命が変更される場合でも、初期原腸胚の場合よりも長い時間が掛かった。
- シュペーマンとマンゴルドの実験 - 原口背唇部の移植実験(1924年)

ドイツのハンス・シュペーマンは、イモリの胚の交換移植実験を行った。原腸胚初期の原口の上部(原口背唇)を切り出し、同じく原腸胚初期の他の胚の外胚葉の表皮になる予定の部分へ移植した。すると頭が2つある幼生ができた。シュペーマンは、これを移植した細胞が周りの細胞に頭部になるよう情報を伝えたと考えた。原口背唇のように、胚のほかの部分に働きかけ、分化を起こさせる部分を形成体(けいせいたい)、あるいは オーガナイザー(organizer)と呼び、その働き(分化を起こさせる働き)を誘導(ゆうどう、induction)と呼ぶ。この実験結果から、原口背唇は近くの外胚葉に働きかけて、神経管を作る働きがあることが分かる。現代では、移植した細胞からタンパク質が分泌され、これが誘導を行っていることがわかっている。


イモリの眼の形成過程は次の順で起こる。
・一次誘導 原口背唇が形成体(一次形成体)として働き、外肺葉から神経管を誘導する。神経管の前方部は脳(のう)に分化し、脳の両側から一対の眼胞(がんぽう)が生じる。さらに眼胞はくぼんで眼杯(がんぱい)となる。
・二次誘導 眼杯が形成体(二次形成体)として働き、表皮から水晶体(すいしょうたい)を誘導する。眼杯は網膜(もうまく)に分化する。
・三次誘導 水晶体が形成体(三次形成体)として働き、表皮から角膜(かくまく)を誘導する。
このように、誘導の連鎖によって器官が作られていく。
- 中胚葉誘導

A: アニマルキャップ
オランダのニューコープは、メキシコサンショウウオを用いた実験により、内胚葉が外肺葉を中胚葉へと誘導することを示した。これを中胚葉誘導()という。

再生
編集体の一部が失われた場合、その部分が再び作り出されることを再生(さいせい、regeneration)と呼ぶ。
例えば、プラナリアは体を切り刻まれても、切り刻まれた部分が元の体に戻る。 プラナリアを切断すると、切断面に未分化の細胞が集まって再生芽という細胞群ができる。 この再生芽が増殖し、頭部側のものは尾部へ、尾部側のものは頭部へ分化していく。 このとき頭部と尾部の方向は切断する前と同じになる。
また、イモリは、手足や尾の一部が失われても、元に戻る。 イモリの手や足を切断すると、切断面の細胞が脱分化して再生芽ができる。 また、イモリの眼の水晶体を除去しても、虹彩の背側の色素細胞から水晶体が再生したりもする。
ヒトも傷や骨折が治るので、ある程度再生する能力を持っているといえる。 近年では、心不全の治療のために、筋肉組織から筋肉細胞を取り出し、培養し、シート状にして心臓に張り付けるなどの再生医療()の研究も進んでいる。
アポトーシス
編集発生などの段階で、ある細胞では、遺伝的にあらかじめ死ぬようにプログラムされている細胞がある。たとえば哺乳類や鳥類の胚では指と指の間に 水かき が始めのころにあるが、この水かきの所の細胞は死んで組織が退化していく。このような、あらかじめ死ぬようにプログラムされた細胞死をプログラム細胞死という。ヒトの手足の指の間の部分も、発生時に水かきのようなものがプログラム細胞死をしている。 カエルの幼生(オタマジャクシ)が変態で尾がなくなるのもプログラム細胞死である。 正常な発生のためにプログラム細胞死は必要なことである。
プログラム細胞死の多くは、まず細胞膜および細胞小器官は正常なまま染色体・DNAだけが凝縮し、それによって細胞膜が変化するなどして細胞が断片化して壊れて死んでいく。このような細胞死をアポトーシス(apoptosis) という。 ヒトやニワトリの手足の指の間の部分の発生時に水かきのプログラム細胞死も、アポトーシスである。オタマジャクシの尾がカエルへの変態で無くなるプログラム細胞死もアポトーシスである。
なお、いっぽう、傷や栄養不足や病原菌などによって細胞が壊されるなどして死んでいくことを壊死(えし)またはネクローシス(necrosis) という。
ニワトリの発生
編集※ 資料集などに書いてある。深入りの必要は無い。
ヒトの発生
編集※ 保健体育などと、ほぼ同内容だが、高校生は教養として、目を通しておく程度には勉強しておくこと。大学で生物系に進学する場合、基礎知識として必要になる。
※ 生物基礎の範囲外 or 発展
編集発展: 細胞融合
編集※ 現代では、基本的に高校『生物基礎』『生物』の範囲外になっている。もし教科書に書いてあったとしても、コラムなどだろう。
発展: クローン動物
編集- テロメア
※ 第一学習社や数研出版の教科書に記述あり。
発展: 幹細胞
編集- ガードンの核移植実験
- ES細胞(胚性幹細胞)
- iPS細胞
- ※ 専門生物の教科書にコラムなどで書いてあったりするが、高校の段階では理解しきれないので、深入りの必要は無い。
脚注
編集参考文献
編集- 田中隆荘ほか『高等学校生物I』第一学習社、2004年2月10日発行、pp.66-109
- 『NHK高校講座 生物』第9-15回
- 生物学用語辞典 - Weblio 学問