高等学校生物/生物I/環境と動物の反応
高等学校生物 > 生物I > 環境と動物の反応
刺激の受容と反応
編集刺激の受容
編集生物に作用して反応を起こさせる要因を刺激()と呼ぶ。 眼や耳などの、刺激を受け取る器官を受容器(じゅようき、receptor)という。生物が刺激に対して活動を起こすことを反応(reaction)と呼ぶ。 反応は筋肉や腺などの効果器(こうかき、effector)で引き起こされる。効果器のことを作動体ともいう。そして受容器と効果器との間は神経系で結ばれている。
受容から反応まで、次のような順序である。
- 刺激 → 受容器 → 中枢 → 効果器 → 反応
また、刺激を受けた感覚細胞が活動状態となることを興奮(こうふん)という。興奮の正体は、細胞膜の電気的な変化である。
受容器
編集受容器はそれぞれ受容する刺激が決まっており、受容できる刺激を適刺激(てきしげき、adequate stimuli)と呼ぶ。 ヒトの五感と受容器と適刺激は次の表のようになっている。
| 感覚 | 受容器 | 適刺激 |
|---|---|---|
| 視覚 | 眼 | 光(可視光線) |
| 聴覚 | 耳 | 音(空気の振動) |
| 嗅覚 | 鼻 | 気体の化学物質 |
| 味覚 | 舌 | 液体の化学物質 |
| 触覚 | 皮膚 | 圧力 |
感覚細胞は一定以上の強さの適刺激を受けないと興奮しない。興奮するための刺激の最小値のことを閾値(いきち、threshold)という。
様々な受容器
編集眼の構造と働き
編集
眼のように光を受容する器官を'視覚器(optic organ)'と呼び、 光の感覚を視覚(vision)と呼ぶ。 ヒトの眼はカメラとよく似た仕組みになっており、このヒトの眼の仕組みをカメラ眼という。 眼は、水晶体(すいしょうたい、lens)で光を屈折し、網膜(もうまく、retina)に像を上下左右逆に結ぶ。カメラに例えると、水晶体がレンズに相当し、網膜がフィルムに相当する。
ヒトの眼での遠近のピント調整は、カメラでいうレンズに相当する水晶体の厚さをかえることで遠近のピントを調整している。機械式カメラとは違って、レンズの前後移動に相当するような仕組みは無い。
ヒトの眼は、前部の表面に角膜(cornea)があり、その内側に瞳孔(pupil)と虹彩(iris)があり、さらにその内側に水晶体(lens)とチン小帯(Zonule of Zinn)と毛様体(ciliary body)がある。 内部には球形のガラス体(Vitreous humour)があり、それを囲むように網膜(retina)がある。
網膜の盲斑(もうはん、blind spot)からは視神経が伸びている。盲斑には視細胞(しさいぼう、visual cell)が無く、そのため、盲斑に像が写っても見えない。盲斑のことを盲点(もうてん)ともいう。
光は視細胞で電気信号にかえられ、その電気信号が視神経を通り脳(主に大脳)へ送られ、視覚が発生する。
網膜の視細胞(visual cell)には二種類の視細胞があり、明暗を感じるかん体細胞(かんたいさいぼう、桿体細胞、rod cell)と、色を感じる錐体細胞(すいたいさいぼう、cone cell)がある。
かん体細胞は明暗のみを区別し、色は区別しない。視細胞には光を吸収する物質の視物質があり、吸収によって、その細胞の特性が変化することから、それぞれの視細胞で光あるいは色などを感じている。
ヒトやサルの錐体細胞では、三原色の赤・青・緑の区別をしており、三種類の錐体細胞(青錐体細胞、赤錐体細胞、緑錐体細胞)でこれらの色を区別しており、色覚が生じている。三種類の錐体細胞は、光の波長によって感度が異なり、それぞれ420nm(青)、530nm(緑)、560nm(赤)を中心に吸収する。このような仕組みで色覚が生じている。なお、緑と赤が近い。 こららの三種類の錐体細胞では、それぞれの色に対応する視物質のフォトプシンがふくまれており、その色の光を良く吸収する。そのため三種類の錐体細胞の色の感度が異なる。
錐体細胞は、網膜の中央部の黄斑に多く分布する。
明るいところで、錐体細胞は、よく働く。弱い光では錐体細胞は反応しない。このため暗いところでは色を区別できない。
錐体細胞の色の光を吸収する色素は、光を吸収すると一時的に分解する。この分解を視細胞が感じ取っており、色覚を生じている。
白色の光は、赤・青・緑のすべての色をふくんでいる光であり、白色光があたると三種類の錐体細胞が三種類とも興奮する。
動物によっては、錐体細胞の種類の数が異なり、そのためヒトとは異なる色の世界を見ている動物も多い。
かん体細胞にはロドプシンという感光する物質が含まれている。ロドプシンの色が紅色なので視紅(しこう)ともいう。ロドプシンに光が当たると、レチナールとオプシン(タンパク質の一種)に分解される。この際、かん体細胞での細胞膜のイオンの透過性が変化し、そのため細胞が興奮する。
ロドプシンはビタミンAから作られる。そのためビタミンAが不足するとロドプシンが不足するので、暗いところで物が見えなくなる夜盲症(やもうしょう)になる。
- ※ ビタミンについてはwikibooks『高等学校化学I/その他』でも、他のビタミン(ビタミンCなど)についての説明がある。目とはあまり関係ないので、この高校生物のページでは深入りしない。
暗いところから明るいところになったとき、
視覚器が次第になれてくることを明順応(めいじゅんのう、light adaptation)といい、
その逆を暗順応(あんじゅんのう、dark adaptation)という。
明順応の仕組み
- 暗いところから急に明るいところに出ると、視物質のロドプシンが急に分解され、そのため かん体細胞 が急激に興奮する。ロドプシンが分解されるにつれ感度が低下するので、明るさに慣れてくる。(※ 範囲外: 明順応の実態は、単に、後述の「暗順応」が終了しただけ[1]だと考えられている。)
暗順応の仕組み
- ロドプシンが分解されないので、ロドプシンが蓄積される。そのため感度が上がり、暗くても光が見えやすくなってくる。

眼は、明暗を虹彩にある瞳孔を拡大縮小することで調節している。明るい所では瞳孔は小さくなる。暗いところでは瞳孔が大きくなる。 また、遠近を水晶体を厚くしたり薄くしたりすることで調節している。
- チン小帯
遠くを見るとき、水晶体はチン小帯に引っ張られてうすくなり、このため焦点距離が長くなり(屈折率は小さくなり)、遠くの物が網膜上に像を結ぶ。チン小帯は毛様体の筋肉に引っ張られて調節される。
いっぽう、近くを見るとき、チン小帯がゆるみ、水晶体は自らの弾性で厚くなる。このため屈折率が大きくなり焦点距離が短くなり、ちかくの物が網膜上に像を結ぶ。
耳の構造と働き
編集
外耳には耳殻(pinna)、外耳道(ear canal)があり、 中耳には鼓膜(こまく、eardrum)、耳小骨(じしょうこつ、ossicle)、耳管(ユースタキー管, Eustachian tube)があり、 内耳には半規管(はんきかん、semicircular canals)、前庭(ぜんてい、vestibule)、うずまき管(cochlear duct)、聴神経(auditory nerve)がある。ツチ骨(槌骨)とキヌタ骨とアブミ骨をまとめて耳小骨(じしょうこつ)という。
- 聴覚
ヒトの耳のように音を受容する器官を聴覚器(ちょうかくき、auditory organ)と呼び、 音の感覚を聴覚(ちょうかく、hearing)と呼ぶ。
ヒトの耳は、外耳(がいじ、outer ear)、中耳(ちゅうじ、middle ear)、内耳(ないじ、inner ear)の3つの部分からなる。音波を受容する聴細胞(ちょうさいぼう)は内耳にある。
音は空気の振動であり、空気の波である。音の波を、音波(おんぱ)という。音は、外耳の耳殻で集められ、外耳道を通る。
音は中耳にある鼓膜(こまく、eardrum)を振動させ、耳小骨(じしょうこつ、ossicle)によって振動が増幅される。
振動は内耳にあるうずまき管(cochlea)を満たすリンパ液に伝わる。
リンパ液の振動は、うずまき管内の基底膜を振動させ、基底膜のコルチ器(Corti's organ)と呼ばれる部分の聴細胞(ちょうさいぼう)の感覚毛を変形させ、聴細胞が興奮する。詳しく言うと、リンパ液の振動によって、コルチ器の聴細胞の感覚毛が、その上をおおっているおおい膜(tectorial membrane)と接触し、その結果、コルチ器の聴細胞が興奮して、最終的に聴覚が生じる。
聴細胞の興奮は、聴神経(ちょうしんけい)によって大脳に伝わって、こうして聴覚(ちょうかく)が発生する。



- 平衡覚

また、耳は聴覚のほかに、からだの傾きなどを感じる平衡覚(へいこうかく)を感じる。 からだの姿勢・動作を知る感覚を、平衡覚(sensation of equilibrium)と呼ぶ。
平衡覚は、前庭(ぜんてい、vestibule)と半規管(はんきかん、semicircular canals)によって感じる。内耳に、前庭と半規管がある。 前庭では、感覚毛(vibrissa)を持った感覚細胞があり、この上に耳石(じせき、otoconium)という石灰質(炭酸カルシウム)の粒子が乗っている。体が傾くと、前庭では耳石が動き、感覚毛を持った感覚細胞が刺激として受け取るので、こうして体の傾きを感じる。耳石のことを平衡砂(へいこうさ)あるいは平衡石(へいこうせき)ともいう。


また、体が回転すると、半規管ではリンパ液がうごき、それを感覚毛をもった有毛の感覚細胞が刺激として受け取るので、こうして体の回転を感じる。
体の回転を止めても、感覚では回りつづけるような感じがする現象、いわゆる「目が回る」現象のある理由は、体の回転を止めてもリンパ液は慣性によって、しばらく流れ続けているからである。
半規管では3個の半円状の管があり、この3個の半規管は、それぞれ直交して約90度をなす配置になっている。この3つの半規管によって、それぞれ前後・左右・水平の3方向の平衡感覚を区別している。 半規管の一方の根元にはふくらんだ部分があり、そこの内部に有毛の感覚細胞がある。
- 参考: 音の高低の識別

基底膜の振動する箇所が、音の高低によって違う。なお、音の高低の正体とは、音波の振動数の違いであり、振動数が大きいほど音も高い。 振動数が大きい音ほど、うずまき管の入口ちかくのを振動させ、つまり鼓膜に近いがわが振動する。 いっぽう、振動数が小さい音ほど、うずまき管の奥を振動させる。
- 超音波
ヒトは20000Hz(ヘルツ)以上の音を聞き取ることが出来ない。Hzとは1秒間あたりの振動数。つまり20000Hzとは1秒間につき2万回の振動ということ。ヒトが聞き取れないほどに高い音波のことを超音波(ちょうおんぱ、英:ultrasonic)という。コウモリなど、いくつかの動物には、超音波を聞き取れるものがいる。コウモリは飛びながら超音波を発し、反射して帰ってきた超音波を感じることができるので、これによって夜間などでも周囲の状況を知ることができる。
鼻の構造と働き
編集
鼻のように気体の化学物質を受容する器官を嗅覚器(きゅうかくき、olfactory organ)と呼び、 その感覚を嗅覚(きゅうかく、olfaction)と呼ぶ。
鼻には、入口の鼻孔(nostril)、その奥の広い鼻腔(nasal cavity)、鼻腔の上部の嗅上皮(きゅうじょうひ、olfactory epithelia)がある。 嗅上皮には、嗅細胞(きゅうさいぼう、olfactory cell)があり、表面の粘液層に繊毛をだし、粘液に溶け出した化学物質を嗅細胞の受容体が受容して興奮する。受容体に種類があり、種類ごとに結合できる物質がちがうので、それによって、においを区別できる。受容体の結合によってイオンチャネルが開き、電位が変化して、興奮する。嗅細胞の興奮が嗅神経によって脳へ伝えられていき、脳で嗅覚として認識する。
舌の構造と働き
編集舌のように液体の化学物質を受容する器官を味覚器(みかくき、Gustatory organ)と呼び、 その感覚を味覚(みかく、gustation)と呼ぶ。
舌の表面には、舌乳頭(ぜつにゅうとう)と呼ばれるつぶつぶが多数あり、 舌乳頭には、味覚芽(みかくが、gustatory bud)と呼ばれる受容器が多数あり、この味覚芽に受容体がある。 味覚芽には、味孔()と呼ばれる孔の奥に味細胞(みさいぼう、gustatory cell)があり、この味細胞の細胞膜にタンパク質でできた受容体があり、 その味細胞の受容体が水などに溶け出した化学物質を受容する。
ヒトの味覚には、甘味(あまみ)、塩味(しおみ)、苦味(にがみ)、酸味(さんみ)、うま味(うまみ) の5つがある。コンブにふくまれるグルタミン酸ナトリウムなどが、うま味をひきおこす物質である。カツオブシのイノシン酸ナトリウムも、うまみをひきおこす。 日本人の池田菊苗が、グルタミン酸ナトリウムによる、うま味を発見した。なお、池田の弟子の木霊新太郎がカツオブシのイノシン酸ナトリウムのうま味を発見した。
特定企業の商品だが「味の素」の主成分が、グルタミン酸ナトリウムである。
グルタミン酸は核酸の主成分であり、イノシン酸は核酸の主成分である。
塩味など、水などに溶けた化学物質が受容体に結合すると、チャネルが開き、電位が変化してシナプスから神経伝達物質を放出し、味神経を興奮させ、興奮が脳へ伝えられていき、脳で味覚を認識する。
私たちが甘みやうまみを「おいしい」と感じるのは、その感覚を起こす物質が生きるのに必要な場合が多いからである。たとえば甘みなら、砂糖などの等分が含まれており、エネルギーの摂取に役立つ。うま味の物質はタンパク質やアミノ酸などの場合が多く、肉体を構成するのに必要な物質である。逆に、苦味を「まずい」味だと感じるのは、それが危険な物質である場合が多いからである。酸味は、腐敗物にふくまれる場合があり、そのため、注意が必要な味として感じているだろう、などと思われている。
トウガラシにふくまれる化学物質のカプサイシンの辛み(からみ)は、痛覚を刺激しており、触角に近い「痛み」の感覚であり、純粋な味覚ではない。ところが、このような辛みを、脳は「味」として認識することから、どうやら味覚と触覚の感覚は、似たような受容の仕組みを持っているらしい、とも言われてる。まだ学者たちが辛みについては研究中なので、高校は深入りする必要は無い。
皮膚の構造と働き
編集
皮膚のように接触の刺激を受容する器官を触覚器()と呼び、 その感覚を触覚()と呼ぶ。 また、皮膚は触覚のほかに温覚、冷覚、痛覚を感じる。
触覚を感じるのはメルケル小体()やマイスナー小体(Meissner corpuscle)やパチーニ小体(pacinian corpuscle)(触点)、 温覚(sensation of warm)を感じるのはルッフィーニ小体()(温点)、 冷覚(cold sensation)を感じるのはクラウゼ小体()(冷点)、 痛覚()を感じるのは痛点(つうてん)という神経の自由末端である。
様々な効果器
編集筋肉の構造と働き
編集

筋肉の内、骨格筋(きんせんい、skeletal muscle)は、自分の意志で動かすことができる。 骨格筋には、屈筋(くっきん、flexor muscle)と伸筋(しんきん、protractor muscle)があり、これによって腕や脚を曲げたり伸ばしたりできる。
骨格筋の筋繊維(きんせんい、muscle fiber)は多核の細胞であり、 筋繊維の中には多数の筋原線維(きんげんせんい、myofibril)が束になっている。 つまり、筋原繊維の束(たば)が筋繊維である。筋繊維の束が骨格筋などのそれぞれの筋肉である。
筋原繊維は、光学顕微鏡で観察すると、明るい明帯(めいたい)と、暗い暗帯(あんたい)とが、交互に並んでいる。明るく見える部分は明帯(めいたい、light bands)といい、
暗く見える部分は暗帯(あんたい、dark bands)という。
明帯の中央にある仕切りをZ膜()という。
Z膜とZ膜との間をサルコメア(筋節、sarcomere)といい、このサルコメアが筋収縮の単位がある。
筋繊維は細いアクチンフィラメントと、太いほうがミオシンフィラメントで、できている。 アクチンが明帯であり、ミオシンが暗帯である。
- ※ なお、アクチンとミオシンは、細胞骨格にも関わる成分である。『高等学校生物/生物I/細胞の構造とはたらき#発展:_細胞骨格』を参照せよ。
- 細胞骨格の「アクチンフィラメント」と、筋肉の「アクチンフィラメント」とは、同種・同類のものだと考えられている。(数研出版・第一学習社などの見解)
この骨格筋の縞模様のことを横紋(おうもん)ともいい、骨格筋には横紋が見られるので骨格筋のことを横紋筋(おうもんきん)ともいう。
ミオシンはATP分解酵素を持っており、運動のためにATPを分解してADPにする。筋肉は、このATPのエネルギーを利用して、力を出している。
なお、一般に、ミオシンのような運動を発生させるタンパク質のことを「モータータンパク質」という。
(※ 専門『生物』の範囲外)余談だが、筋肉組織だけでなく、微小管上を移動するキネシンとダイニンもモータータンパク質であることが知られている。なお、キネシンとダイニンもそれぞれATPを分解する部位を持つ。(※ 一部の教科書で紹介。)
- 滑り説

筋収縮では、ミオシンフィラメントの間にアクチンフィラメントが滑り込む。この説を滑り説(すべりせつ、sliding filament model)という。
比喩として、よくアクチンが鉄道などのレールにたとえられ、ミオシンのほうがレールの上を移動する何らかの移動体などに(ミオシンが)例えられる(啓林館の教科書にもある比喩)。
なお、余談だが、植物の原形質流動でも、ミオシンとアクチンとの何らかの相互作用が起きている、と考えられている(※ 参考文献: 第一学習社の専門『生物』)。また、アメーバの運動は、アクチンによるものである(※ 参考文献: 数研出版の専門『生物』)。
筋原繊維は、筋小胞体に囲まれている。
神経の刺激によって活動電位が発生したさい、筋小胞体からCa2+が放出される。 このCa2+がの作用で、アクチンフィラメントにあるトロポニンと結合し、アクチンフィラメントに付着しているトロポミオシンの構造が変化することで、トロポミオシンによってさえぎられていたアクチンのミオシン結合部位が露出し、アクチンフィラメントがミオシンと作用できるようになり、よって筋収縮が起きる。こうしてサルコメアが収縮することで、筋収縮が起きている。
- 筋収縮

カエルのふくらはぎの骨格筋にへの電気刺激の収縮量の測定実験(キモグラフを用いる)で、つぎの段落で説明する単収縮・強縮のしくみが事が明らかになってる。

カエルなどの実験動物の骨格筋に運動神経を付けたまま取り出したものを、神経筋標本という。
実験動物の座骨神経(ざこつ しんけい)のついたままの神経筋標本に、1回の短い電気刺激を与えると、収縮したのち、すぐ(0.1秒ほど)に弛緩(しかん)する。この1回の電気刺激で起こる1回の収縮を単収縮(たんしゅうしゅく、twitch)という。単収縮のことを、れん縮(れんしゅく, spasm)ともいう。

- ※ 高校の理科の検定教科書では、坐骨(ざこつ)の表記は「座骨」です。大学教科書などでは「坐骨」(ざこつ)の表記が使われる場合もあります。
筋肉が弛緩する前に次の電気刺激を行うことを繰り返しつづけると、持続的で強い収縮を行う。この強い収縮を強縮(きょうしゅく、tetanus)という。 動物の骨格筋の運動での収縮は、普通は、強縮である場合が多い。
強縮でも、刺激の頻度が低ければ(1秒間に15回の割合)、測定される波形は、単収縮が重なり合ったようなギザギザした形の不完全強縮になる。刺激の頻度がじゅうぶんに多ければ(1秒間に30回の割合)、完全強縮になる。
筋繊維はニューロンによって制御されているため、神経線維の「全か無かの法則」と同様、1本の筋繊維も、刺激の強さが収縮を起こせる一定値(閾値)以上の強さの刺激があれば筋繊維は収縮し、刺激が一定の強さに届かなければ収縮しない。
閾値は筋繊維の一本一本ごとに違う。
- (※編集者への注意 ここに運動単位を図示してください。)
一本の運動ニューロン(motor neuron)が枝分かれして多くの筋繊維を制御する。この一本の運動ニューロンによって管理されている筋肉を、それを管理する運動ニューロンとまとめて、運動単位(うんどうたんい)という。
筋肉が収縮するさいの直接のエネルギー源はATPである。筋肉の収縮は、ATPを消費して、ATPがADPに変化する。ミオシンの頭部にATP結合部位があり、このミオシン頭部がATP分解酵素としても働き、こうしてATPを分解することで筋収縮のエネルギーを得ている。
急激な運動などで、呼吸や解糖によるATP合成が追いつかない場合は、筋肉にたくわえられているクレアチンリン酸(phosphocreatine)を用いて、ATPを合成する。

クレアチンリン酸は、ATPと同様に高エネルギーリン酸結合を持っている。
休息時などでATPが十分にあるときに、ATPのエネルギーを用いて、クレアチンからクレアチンリン酸が合成され、クレアチンリン酸が貯蔵され、エネルギーを蓄えている。
筋線維を取り出し、それに光またはX線を当てると、光の回折(かいせつ)が見える。その回折光のパターンは、筋線維の横紋の周期に由来していると考えられている[3]。
- ※ 物理学でいう光の「回折」については『高等学校物理/物理I/波/波の性質#回折』を参照せよ。
これを用いて、筋節(きんせつ)の長さを測定できる。筋節とは、横紋の1周期ぶんの長さのことで、普通は Z帯から次のZ帯までの長さ を採用する[4]。
その他の効果器
編集- 発電器官
シビレエイやデンキウナギなどが発電器官をもつ生物には発電器官がある。 発電器官は筋肉が変化した発電板()が多数重なってできている。 発電版には片側に神経が分布している。 発電版は普段は外側が+で内側が-であり、発電器官を電流が流れることはないが、 興奮時は神経が分布している側の電位が逆になり、発電板が直列につながることで高電圧を生じる。 シビレエイは50~60V、デンキウナギは800Vの起電力が測定される。
- 発光器官
ホタルは腹部に発光器官を持つ。 発光器官をもつ生物には、ホタル、ホタルイカ、オキアミなどが挙げられる。 ホタルの発光器官は発光細胞層()と反射細胞層()からなる。 発光細胞層から発光物質を分泌し、気管から取り込んだ酸素と反応させ発光させ、反射細胞層で光を外側に反射する。 ウミホタルは口の近くの発光腺から発光物質を分泌する。 この物質が体外で酸化し発光する。
ホタルは、雌と雄とが出会う手段として、自己の発光を利用している。
・ルシフェリンとルシフェラーゼ
ホタルの尻尾にある器官に、発光物質のルシフェリンがある。ルシフェリンが、酵素のルシフェラーゼが触媒として、ATPと酸素O2と反応して、酸化ルシフェリン(オキシルシフェリン)になる。この反応に伴って、発光が起こる。
- ルシフェリン + ATP → 酸化ルシフェリン + 光
- (触媒:ルシフェラーゼ)
・ルシフェリンの応用(おぼえなくて良い。範囲外。参考。)
よってATPの量の測定手段として、ルシフェリンと蛍光光度計を用いることにより、ATP量が測定できる。微生物量測定などのバイオテクノロジーにもルシフェリンが利用されている。また、遺伝子組み換え実験などでも、暗闇で光らせられるので、目的の細胞を見分けるためのマーカーとしても利用されている。
ATP量の測定については、反応する前のルシフェリンとルシフェラーゼの量を、一定にしておけば、ATPの量によって発光の強さが変わるからである。ところで、ほとんどの細菌は体内にATPをもつから、ルシフェリンを用いて、細菌の量を測定できる。つまり、微生物による汚染の度合いを測定できる。
遺伝子組み換えについては、ルシフェラーゼをつくる遺伝子を目的の細胞に導入しておくと、ルシフェラーゼの導入された植物は、暗闇で光り輝くので、融合が成功したかどうかを確かめることができる。ルシフェラーゼ遺伝子のように、細胞融合が成功したかどうかを確かめるための遺伝子をマーカーという。
- 色素胞と体色変化
動物の体色が変化する現象を体色変化()と呼ぶ。 体色変化する生物には、ヒラメやカメレオンなどが挙げられる。 メダカの体色変化は、色素胞(しきそほう)と呼ばれる細胞で、内部にある色素果粒()が、神経やホルモンの働きにより、凝縮したり拡散したりすることで起こる。
生物が能動的に音を出すことを発音と呼び、 発音を行う器官を発音器官()と呼ぶ。 ヒトの発音器官は咽頭部の声門(glottis)である。 声門の軟骨の間にある声帯(vocal cord)と呼ばれる部分が、通過する空気によって振動して声が出る。
腺の構造と働き
編集特定の物質を分泌する器官を腺(せん、gland)と呼ぶ。 腺には、体外に分泌する外分泌腺(がいぶんぴせん、exocrine gland)と、体内に分泌する内分泌腺(ないぶんぴせん、endocrine gland)がある。
内分泌腺はホルモンを体内の血流に分泌する。内分泌されたホルモンは血流によって全身に運ばれる。
外分泌腺には、皮膚で汗を分泌する汗腺(かんせん、sweat gland)や、口で唾液を分泌する だ腺(だせん、salivary gland)などがある。 分泌物は腺細胞()で作られ、排出管()を通り分泌される。
神経
編集ニューロンの構造と働き
編集

刺激をある器官から別の器官へ伝える器官系を神経系(しんけいけい、nervous system)と呼ぶ。 神経系はニューロン(neuron)と呼ばれる神経細胞から成り立っている。
ニューロンは、細胞体(さいぼうたいcell body)と、細胞体の周りの多数の樹状突起(じゅじょうとっき、dendrite)と、細胞体から伸びる一本の軸索(じくさく、axon)からなる。
軸索はシュワン細胞(Schwann cell)でできた神経鞘(しんけいしょう、neurilemma)で囲まれており、軸索と神経鞘をあわせて神経線維(nerve fiber)と呼ぶ。 神経線維には髄鞘のある有髄神経線維()と、髄鞘のない無髄神経線維()とがある。 有髄神経線維の髄鞘のないくびれをランビエ絞輪(ランビエこうりん、Ranvier's constriction ring)と呼ぶ。 ニューロンとニューロンの連結部をシナプス(synapse)と呼ぶ。

刺激を受けた細胞が休止状態から活動状態になることを興奮(こうふん、excitation)と呼ぶ。 興奮がニューロンの中を伝わることを伝導(でんどう、conduction)と呼び、 シナプスを介してあるニューロンから別のニューロンへ刺激の情報が伝わることを伝達(でんたつ、transmission)と呼ぶ。
伝導はニューロンの電気的変化で伝えられる。この電気を起こす正体はニューロンの細胞膜にあるイオンポンプやイオンチャネルの働きである。そのため、神経細胞は体液に取り囲まれている。神経での伝導は、金属の電気伝導などとはちがい、ニューロンの興奮の伝導では電気が伝わるのに時間が掛かる。(無髄神経線維を興奮が伝導する速さは1m/秒程度。 有髄神経線維を興奮が伝導する速さは100m/秒程度である。) ニューロンの細胞内は刺激を受ける前、細胞内は負に帯電しており、膜外を基準にすると膜内は -90mV ~ -60mV ( 平均 -70mV ) の負の電位をもっており、このような刺激を受ける前の膜内の負の電位を静止電位(せいしでんい、resting potential)という。
ニューロンの一部に刺激を受けると、一瞬、刺激を受けた場所の電位が変化する。
まず、刺激を受けた直後、刺激を受けた場所の細胞内の電位は一瞬、細胞の内側が外側よりも高い電位になり、細胞内は約+40mVの電位をもつ。その後、すぐ(約1/1000秒)もとの静止電位にもどる。このような電位の変化を活動電位(かつどうでんい、action potential)という。 神経の興奮の正体は、活動電位の発生である。
ニューロンの一部分の興奮は、ニューロン上のとなりの細胞へと伝わっていく。これが伝導(でんどう)である。その結果、興奮は、ニューロン線維の両側へと伝導していき、ニューロンの両端まで伝わっていく。
- イオンチャネルの働き
- ※ 一般的なイオンチャネルについては、すでに『高等学校生物/生物I/細胞の構造とはたらき#受動輸送』で説明してある。
- 本『環境と動物の反応』のページでは、主に神経細胞のイオンチャネルについて説明する。(※ 高3の専門生物の範囲内です。)
神経細胞の活動電位にも、神経細胞の膜表面にあるイオンチャネルとナトリウムポンプが関係している。
1
- 神経細胞でも、静止状態ではナトリウムポンプの働きによって、細胞の内側でナトリウム濃度が高くカリウム濃度が低く、外側ではナトリウム濃度が低くカリウム濃度が高い(他の一般の細胞でのナトリウムポンプの動作と同様の結果の濃度勾配)。
- 一部のカリウムチャネルが刺激の有無に依存せず漏れ出すので(※ 東京書籍・数研出版の検定教科書に記述あり)、つまりプラスの電荷をもったK+が細胞外へ漏れ出すのだから、その結果、静止状態の細胞内は負(-)に帯電している。なお、この現象を「分極」(ぶんきょく)と呼んでいる(※ 東京書籍に記述あり)。
- ※ とりあえず、神経細胞内が初期状態では負に帯電している事が分かればいいだろう。
2
- 神経細胞のナトリウムチャネルは普段は閉じているが、電位が上昇すると、ナトリウムチャネルが開き、細胞内にナトリウムチャネルが流入する。その結果、細胞内の電位が上がり、今までの負(-)の電位から一瞬、正(+)になる。なお、この現象を「脱分極」という(※ 東京書籍に記述あり)。(図「静止電位と活動電位」の、○2の状態)
3
- 上記のように細胞内の電位が正になり膜内外の電位は逆転した結果、ナトリウムチャネルは閉じる。
4
- そして細胞内に入ったナトリウムは、(ナトリウム「チャネル」ではなく)ナトリウムポンプによって外に出されていく。この際、ナトリムポンプの働きによってカリウム(K+)が細胞内に取り込まれていく。
- そして、イオンの分布が、もとにもどる。
- 全か無かの法則

1本のニューロンは、刺激の強さが一定値より弱いと興奮しない。この、さかいめの一定値を閾値(いきち)という。閾値以上だとニューロンは興奮し、その興奮の大きさは刺激の強さによらず一定であり、活動電位の大きさは一定である。 ニューロンは、刺激に対して、興奮するか、興奮しないか、のどちらか2通りだけである。 ニューロンの、このような反応の現れ方を全か無かの法則(ぜんかむかのほうそく、all-or-none law)と呼ぶ。
- 興奮の伝導

このようにして軸索のある箇所に活動電位が起こると、興奮部と隣接する静止部の間に電流が生じ、その電流を活動電流(かつどうでんりゅう、action current)という。活動電流によって隣(となり)の静止部に興奮が起き、さらに、その興奮によって、そのまた隣の静止部に興奮が起き・・・、というように活動電流によって次々と興奮が伝わっていく。 これを興奮の伝導(でんどう、conduction)という。
興奮をした直後の部位は、しばらく興奮しない状態になる。しくみは、イオンチャネルがしばらく不活性になるからである。この興奮直後の部位の刺激に応答しない時期のことを、不応期(ふのうき)という。
このため、刺激を受けた場所には興奮は戻らず、刺激は静止している側へと伝わっていく。
- 興奮の伝導速度

興奮を伝わる速度は、無髄神経繊維よりも、有髄神経線維のほうが、興奮の伝わる速度が速い。 この理由は、有髄神経繊維では髄鞘(ずいしょう)は電気を通しにくい電気絶縁体であり、活動電流がランビエ絞輪(ランビエこうりん)から隣のランビエ絞輪へと飛び飛びに伝わるためである。このように有髄神経線維にて、興奮がとびとびに絞輪から次の絞輪へと伝導する現象のことを跳躍伝導(ちょうやくでんどう、 saltatory conduction)という。
無髄神経繊維を興奮が伝導する速さは1m/秒程度で、 有髄神経繊維を興奮が伝導する速さは100m/秒程度である。
- (※ なお、解剖学では人体各部の繊維のことを「線維」とも書く。高校生物の検定教科書(2015年に確認。)では「繊維」を用いている。)
| 神経繊維 | 伝導速度 (m/秒) |
太さ (μm) |
温度 (℃) |
|---|---|---|---|
| イカ (無髄) | 25 | 600 | 18 |
| カニ (無髄) | 4 | 30 | 21 |
| カエル (有髄) | 30 | 15 | 22 |
| ネコ (有髄) | 100 | 15 | 37 |
| ネコ (無髄) | 1 | 0.8 | 37 |
文献によって、伝導速度や太さや温度などの細かな数値は、少し違う。なので、細かい数値は、おぼえなくて良い。たとえばネコの場合、文献によって、伝導速度が120だったり110だったり100だったりする。だいたいの数値を把握すればよい。
※参考文献(伝導速度の数値の出典)
- 上記の表中のイカの数値の出典
- (検定教科書)吉里勝利など『高等学校生物』第一学習社、平成24年検定済、平成26年2月10日、p.285・ よりイカの値を参考・引用。
その他、各社の教科書や参考書などを参考文献・引用文献にした。
- (検定教科書)嶋田正和など『生物』数研出版、平成26年1月10日発行
- 水野丈夫など『理解しやすい生物I・II』文英堂、2004年版
- 鈴木考仁など『チャート式シリーズ新生物 生物基礎・生物』数研出版、平成26年4月1日発行
軸策が太いほど、伝導速度が速い。また、温度が40℃未満なら、温度が高いほど、伝導速度が速い。40℃以上に温度が高くなると、伝導しにくくなる。
イカやミミズは、太い神経軸策(巨大神経軸策)を持っており、そのぶん、興奮が伝わる速度も速い。逃げるさいなど、巨大軸策のおかげで信号が早く伝わるので行動の開始が早く、生存に有利だったと考えられている。

ふつう、神経は多数の軸策からなっている。
刺激が大きいほど、神経細胞の興奮の発生頻度が多くなる。なぜなら刺激が強いほど、個々のニューロンでの興奮の頻度も増え、また、多くの感覚細胞が反応することでニューロンも多数が反応するからである。
脳で感じる興奮の大きさの感覚の正体は、神経細胞から伝えられた興奮の発生頻度である。興奮の頻度が高いほど、脳で感じる興奮が大きくなる。
シナプスでの伝達
編集
一つの軸索の先端と、他の神経細胞または筋肉などの効果器との間の部分をシナプスという。神経と筋肉との間のこともシナプスという。一つの神経の信号は、シナプスを経て、つぎの神経または効果器へと伝わる。
シナプスには、小さな隙間(すきま、かんげき)があり、シナプス間隙(シナプスかんげき)という。
シナプスから次のニューロンへと信号を伝える方法は、化学物質の分泌(ぶんぴ、ぶんぴつ)による。そのシナプスでの分泌物を神経伝達物質(しんけいでんたつぶっしつ、neurotransmitter)といい、軸索の末端から分泌される。神経伝達物質には、ノルアドレナリンやアセチルコリン、セロトニン(serotonin)、γアミノ酪酸(ガンマアミノらくさん)、ドーパミン(dopamine)などがある。
交感神経の末端からはノルアドレナリンが分泌される。副交感神経の末端からはアセチルコリンが分泌される。筋肉を動かす神経である運動神経の末端からはアセチルコリンが分泌される。
軸索の末端の内部には、つぶ状のシナプス小胞(シナプスしょうほう、synaptic vesicle)という物質があり、このシナプス小胞に伝達物質が含まれている。シナプスに興奮が伝わるとシナプス小胞から、アセチルコリン(acetylcholine)、ノルアドレナリン(noradrenaline)などの神経伝達物質(neurotransmitter)が分泌されることで、となりの細胞に興奮が伝えられる。
軸索の末端に、電位に依存するカルシウムチャネル( Ca2+チャネル )があり、このCa2+チャネルに活動電位が到達することで、このチャネルが開き、Ca2+が軸策末端の細胞内に流入する。このCa2+の流入によって、シナプス小胞の膜が 軸策末端の膜(シナプス前膜) と融合し、神経伝達物質がシナプス間隙に放出される。 シナプスのうち、放出側の細胞のほうをシナプス前細胞(シナプスぜんさいぼう)といい、その放出側のシナプス前細胞の細胞膜を、シナプス前膜(シナプスせんまく)という。シンプスのうち、受け取り側の細胞のほうをシナプス後細胞(シナプスこうさいぼう)といい、そのシナプス後細胞の細胞膜をシナプス後膜(シナプスこうまく)という。
受取り側の、となりの細胞の細胞膜には、伝達物質の受容体があり、さらに、その受容体によって働きの変わるイオンチャネルがある。(受容体がイオンチャネルを兼ね備えている場合もあるし(イオンチャネル型受容体)、受容体とイオンチャネルがそれぞれ存在する場合もある。 ※ 高校の範囲外だろう。)
さて、伝達物質に依存するイオンチャネルが、受け取り側の細胞膜に存在している。伝達物質依存性のイオンチャネルが、伝達物質と受容体との結合によって働いて、興奮についての信号がとなりの細胞に伝わる。シナプスでの興奮が一方向( シナプス → となりの細胞 )に伝達され、信号は逆流はしない。
シナプスを介してある軸索から、となりの細胞へ興奮についての情報が伝わることを伝達(でんたつ)と呼ぶ。シナプスから出る化学物質によって、興奮の情報は伝達される。
シナプスで放出される神経伝達物質には、興奮をさせる興奮性の物質と、興奮をさせにくくする抑制性の物質とがある。興奮性の物質にはアセチルコリンやノルアドレナリンがある。抑制性の物質には、γアミノ酪酸(ガンマアミノらくさん、GABA)やグリシンがある。
- 同じ神経伝達物質でも、受け取り側の神経細胞によって、興奮性にも抑制性にも、なりうる。たとえば、さきほどアセチルコリンを「興奮性」と言ったが、じつは心臓の迷走神経(めいそうしんけい)などではアセチルコリンが抑制性に働く場合もある。
さて、興奮性の神経伝達の場合では、Na+チャネルが開き、Na+が細胞内に流入して、活動電位が生じる。シナプスに限らず、神経細胞の興奮は、ナトリウムイオンの神経細胞内への流入によって起きている。いっぽう、抑制性の神経伝達物質の場合は、Cl-チャネル(読み:「クロライドチャネル」)が開き、Cl-が細胞内に流入する。 これらのイオンチャネルの働きによって、受け取り側の細胞での膜電位が変わるので、膜電位の高低によって、興奮や抑制の、コントロールが行われている。
Na+チャネルが開けば膜電位は高まり、膜電位が高まれば、受け取り側の細胞は興奮をする。 いっぽう、Cl-チャネルが開けば膜電位は下がり、膜電位が低ければ、受け取り側の細胞は抑制される。 Na+チャネルとCl-チャネルの両方が開けば、膜電位の高低が打ち消しあう。
しだいに神経伝達物質は、再吸収されたり、あるいは酵素(コリンエステラーゼなど)によって分解されたりするので、興奮や抑制は、しだいに終わっていく。そして、次に来る信号が伝達可能になる。
- (※ 範囲外: )アセチルコリンの化学式はエステル構造である[6]。エステルの意味については『高等学校化学I/脂肪族化合物/エステル』を参照のこと。
- ※ 「コリンエステラーゼ」という名前からは まぎらわしいが、コリンエステラーゼは、(コリンをエステル化するのではなく、)アセチルコリンを分解してエステルと酢酸に分解する酵素である。
興奮性の伝達物質を放出するシナプスを興奮性シナプス(excitatory synapse)といい、いっぽう、抑制性の伝達物質を放出するシナプスを抑制性シナプス(inhibitory synapse)という。
シナプスの後膜の電位のことを後電位(こうでんい)あるいは後膜電位(こうまくでんい)という。興奮性シナプスの後電位のことを興奮性シナプス後電位(EPSP:excitatory postsynaptic potential)という。抑制性シナプスの後電位のことを抑制性シナプス後電位(IPSP:inhibitory postsynaptic potential)という。
シナプスで情報がシナプス前細胞からシナプス後細胞に伝わるのに、約1ミリ秒~2ミリ秒がかかり、この遅れ(おくれ)のことをシナプス遅延(シナプスちえん)という。
- 発展:コカインとドーパミン
- 参考:毒ガスのサリン
神経毒のサリンは、アセチルコリンの分解を行う酵素(コリンエステラーゼ)の働きを、さまたげる。(数研の(チャート式だけでなく)専門生物の検定教科書にも書いてある。)
神経系の種類
編集
- (※編集者へ注意 ここにヒドラやプラナリアやミミズや昆虫(バッタあるいはハチなど)の神経系の図を追加してください。)
神経系の種類には、神経細胞(ニューロン)が体全体に散在し網目状に連絡している散在神経系(diffuse nervous system)と、脳・脊髄などに神経細胞(ニューロン)のあつまった集中神経系(concentrated nervous system)がある。
脳・脊髄・神経節などをまとめて中枢神経系(ちゅうすうしんけいけい、central nervous system)という。集中神経系の動物の神経のうち、中枢神経以外の部分の神経を末梢神経系(まっしょうしんけいけい)という。
散在神経系をもつ生物にはイソギンチャクやヒドラやクラゲなどがあげられる。
集中神経系は、脊椎動物などにみられる。ミミズやプラナリアの神経は、集中神経系である。バッタ・ハチなど昆虫の神経系は集中神経系である。
脊椎動物の脳の構造と働き
編集


- (※編集者へ注意 ここにタラ(魚類)の脳、カエル(両生類)の脳、ガチョウ(鳥類)の脳の図を追加してください。)
- ※ 教科書には魚類などの脳の図もあるが、実は参考書には、あまり魚類や両生類の脳は書いてない。少なくとも近年のチャート式では、見あたらない。
感覚器で受けた刺激の情報は感覚神経によって脳(のう、brain)へ送られ、 脳はその情報を判断し、 運動神経によって効果器に情報が送られ反応する。
脊椎生物の脳は大脳(だいのう、cerebrum)、間脳(かんのう、diencephalon)、中脳(ちゅうのう、midbrain)、小脳(しょうのう、cerebellum)、延髄(えんずい、medulla oblongata)からなる。 ヒトの脳には約一千億個のニューロンがあり、そのニューロンには数千のシナプスがあり、複雑なネットワークを形作っている。
- 大脳
大脳の構造は、左右の半球に分かれており、それら左右を結ぶ脳梁(のうりょう、corpus callosum)がある。 両半球は表層は大脳皮質(だいのうひしつ、cerebral cortex)でおおわれており、ニューロンの細胞体があつまって灰色をしているため 灰白質(かいはくしつ)という。 内部には大脳髄質(だいのうずいしつ、cerebral medulla)があり、多くの神経線維が通っていて白色をしているため 白質(はくしつ)という。 大脳皮質には、新皮質(しんひしつ、neocortex)と、古皮質(こひしつ)および原皮質(げんひしつ)からなる辺縁皮質(へんえんひしつ)がある。ヒトの大脳では新皮質が発達している。ヒトの古皮質および原皮質は、大脳に囲まれており、そのため内側に古皮質および原皮質が隠れている。
新皮質には視覚・聴覚など感覚の中枢があり( 感覚野(かんかくや、sensory cortex) )、また、運動の中枢があり( 運動野(うんどうや、motor cortex) )、また、記憶・思考・理解などの学習を必要とする精神活動をつかさどる中枢( 連合野(れんごうや、association cortex) がある。 辺縁皮質は、本能などを司る。辺縁皮質にふくまれる海馬(かいば、hippocampus)という部分が記憶を主につかさどる。
- 脳幹
中脳・間脳・延髄を 脳幹(のうかん、brainstem) という。
- 間脳
間脳の位置は中脳と大脳の間に位置し、構造は視床(ししょう、thalamus)と視床下部(ししょうかぶ、hypothalamus)に分かれている。視床下部に自律神経系の中枢があり、体温の調整や内臓の働きを調整している。また、視床下部は脳下垂体(のうかすいたい)とつながっており、ホルモンの分泌を調整しており、血糖値を調整している。視床は大脳への感覚を中継する。
- 中脳
中脳の構造は、間脳の後方、小脳の上方に位置している。 中脳の働きは、間脳と小脳との通路になっている。眼球運動や瞳孔反射の中枢、聴覚反射、姿勢制御などを司る中枢がある。
- 小脳
小脳の構造は、大脳の後下部に位置している。 小脳には、体の平衡、筋肉の運動機能を司る中枢がある。
- 延髄
延髄の構造は、脳の最下部に位置し、脊髄に続いている。 延髄には、呼吸・血液循環(心臓の拍動)・消化などを司る中枢がある。
延髄より下の体の右側は、脳の左側が担当する。延髄より下の体の左側は、脳の左側が担当する。なぜなら、神経が延髄を通るときに、多くの神経で、左右が交差するからである。したがって脳の右側が損傷すると、体の左側が麻痺(まひ)・不随(ふずい)になる。
参考: 血液脳関門(けつえき のうかんもん)
(※未執筆)
脊髄の構造と働き
編集
さて、中学で習うように、生物学の神経分野でいう「反射」とは、たとえば熱いものに手が触れたときには、思わず手を引っ込めるように、意識とは無関係にすばやく行われる反応である。
また、大脳を介さない反応もあり、脊髄がそのような、大脳を介さない反射の中枢になっているので、そのように大脳を介さないで脊髄が中枢になっている反射のことを脊髄反射(せきずいはんしゃ、spinal reflex)という。
脊髄反射には屈筋反射(くっきんはんしゃ、flexor reflex)やしつがい腱反射(しつがいけんはんしゃ、膝蓋腱反射、knee jerk)などの反射がある。
しつがい腱反射とは、ひざの骨のすぐ下を軽く叩くと、足が勝手に跳ね上がる現象のことである。
しつがい腱反射なら、打撃により、ひざ部の筋紡錘が興奮し、その興奮による信号が感覚神経を伝わっていく。
しつがい腱反射に限らず一般に反射のさい、興奮が伝わる経路のことを 反射弓(はんしゃきゅう、reflex arc)と呼ぶ。
しつがい腱反射の場合の反射弓は
- 受容器→感覚神経→反射中枢→運動神経→効果器
である。
反射は大脳を経由しないため無意識で素早く行われる。
しつがい腱反射は、大脳を介さないので、脊髄反射に分類される。
しつがい腱反射では、しつがい腱をたたくと、大腿四頭筋が縮み、膝関節が伸びる反射を起こす。しつがい腱反射での神経中のシナプスの数は、しつがい腱反射では介在ニューロンを経由せず、よってシナプスは1つである。

屈筋反射(くっきんはんしゃ)では、例えば熱いものに触れた時、手を思わず引っ込めるような、屈筋が縮む反射を起こす。屈筋反射での神経中のシナプスの数は、屈筋反射では介在ニューロンを経由するため、シナプスは2つである。
屈筋反射は、大脳を介さないので、脊髄反射に分類される。
その他の反射として、口に物を入れたときの、だ液の分泌も反射である。だ液の反射中枢は延髄にある。暑いときの発汗も反射である。
目の瞳孔が、光を受けると縮小する、瞳孔の縮小も反射である。瞳孔の反射中枢は中脳にある。
- 解剖学的なこと
- 脊髄(せきずい、spinal cord)は、脳の延髄の下方に続き、脊柱(vertebral column)の管内を通る、中枢神経である。
- 脊髄の内側は細胞体でできた灰白質であり、外側は神経繊維でできた白質である。
- 感覚神経は背根(はいこん、dorsal root)と呼ばれる部分を通り、
- 運動神経と自律神経は腹根(ふくこん、ventral root)と呼ばれる部分を通る。
- 受容器で発生した興奮は、感覚神経によって背根を通り、灰白質・白質を通り、大脳へ伝わる。
- そして、大脳から興奮が、白質・灰白質を通り、運動神経によって腹根を通り、効果器へ伝わる。
- 内臓など
内臓の働きや、消化や、体内のホルモンや血糖の調整なども意識とは無関係に行われるが、これらの現象も「反射」であるとして分類されている(※ 検定教科書の範囲)。内臓など、こういった働きを制御している神経のことを自律神経と言うので、「自律神経」が内臓などの「反射」を調節していると言える。
末梢神経系の種類
編集末梢神経系には、脳から伸びる脳神経(cranial nerves)と、脊髄から伸びる脊髄神経(spinal nerves)とがある。 また、末梢神経系は、体の感覚や運動に関する体性神経系(たいせいしんけいけい、somatic nervous system)と、 意思とは無関係に働く自律神経系(じりつしんけいけい、autonomic nervous system)に分けることもできる。 体性神経系には、感覚神経(sensory nerve)と運動神経(motor neuron)がある。 自律神経系には、交感神経(こうかんしんけい、sympathetic nerve)と副交感神経(ふくこうかんしんけい、parasympathetic nerve)がある。交感神経と副交感神経は対抗的に働くことが多い。()
- 神経系の分類 まとめ
(中央) 神経系━┳━中枢神経系━┳━脳 ┃ ┗━脊髄 ┃ ┗━抹しょう神経系━┳━体性神経系━┳━運動神経 ┃ ┗━感覚神経 ┃ ┗━自律神経系━┳━交感神経 ┗━副交感神経
ヒトの脳神経は12対であり、脊髄神経は31対である。
動物の行動
編集動物が生まれつき持っている行動には、走性と本能行動がある。
2022年度以降の新課程の基礎なし科目「生物」に詳細を載せたので、詳しくはリンク先を見てください。
このページでは、下記に概要を示します。
走性
編集走性(そうせい、taxis)には光走性(phototaxis)、化学走性(chemotaxis)、水流走性(Rheotaxis)、重力走性()、電流走性(galvanotaxis)などがあり、 それぞれ光・化学物質・水流・重力・電流など刺激に対して向かっていくことを正の走性といい、いっぽう遠ざかることを負の走性と呼ぶ。 例えば、プラナリアは負の光走性を持っている。 また、メダカは流れに逆らって泳ぐ正の水流走性を持っている。
光への走性を光走性といい、化学物質への走性を化学走性といい、同様に電気に対しては電気走性という。
- プラナリアの光走性
かぎ刺激
編集
動物には、習わなくても、その行動をする、生得的行動(せいとくてき こうどう)がある。
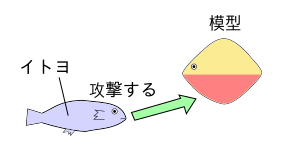
本能行動の研究には、オランダのニコ・ティンバーゲンによるイトヨの研究などがある。 ティンバーゲンは、様々なイトヨの模型を用いて、繁殖期の腹部が赤いイトヨの雄(おす)が、他の雄を攻撃して追い払う本能行動を調べた。
イトヨの雄(おす)は、腹部が赤くないイトヨの模型には反応せず、いっぽうで形が似てなくても下部が赤い模型には攻撃した。
このように、生得的行動を引き起こす刺激をかぎ刺激()と呼ぶ。
つまり、このイトヨの実験では、「イトヨの雄が、他の雄を攻撃する」という生得的行動において、かぎ刺激にあたるものは、腹部の赤い部位である。
イトヨの雌(めす)が卵を産む期間に、イトヨの雄は腹部が赤くなるのである。
-
 イトヨの走性. 赤くない場合.
イトヨの走性. 赤くない場合. -
 イトヨの走性。おおざっぱな形で、赤い腹。
イトヨの走性。おおざっぱな形で、赤い腹。


{kind=link}
学習と知能
編集経験によって得られる行動には、学習行動、知能行動がある。
学習(learning)とは、経験によって行動が変化し、その行動が長く続くことである。 学習には、慣れ、古典的条件付け、試行錯誤、洞察学習、刷り込みなどがある。[7] 例えば、鳥の鳴き方には地鳴きとさえずりがあるが、地鳴きは本能行動であるのに対し、さえずりは学習によるものである。
慣れ
編集軟体動物のアメフラシは、水管に接触刺激をくわえると、えらを引っこめる。しかし、これを繰り返すと、えらを引っこめなくなる。
アメフラシの運動神経は、シナプスを仲介して感覚神経と接続している。
水管への刺激をくりかえしていくと、シナプスで感覚神経末端から放出する神経伝達物質の量が減少するため、えらが反応しなくなるのである。
アメフラシでは、こうして、慣れ(なれ)が起こる。
古典的条件づけ
編集空腹のネズミをカゴに閉じ込め、内部にあるレバーを押すと、餌(えさ)が出て来るような装置を仕掛ける。
すると、はじめのうちは、偶然にレバーを押すことによって、餌を得る。
ところが、しだいにレバーを押す頻度が多くなるので、つまりこの事から、「レバーを押せば餌がでてくること」をネズミが学習したことが分かる。
このような学習を条件づけ(じょうけんづけ)という。
鳥類のすりこみ
編集アヒルやカモなど鳥のひな(雛)は、うまれて初めて見たものを親と認識し、ひなは、親と認識した物体を追いかける。
見せる物が模型などの非生物な物体であっても、ひなは、その初めて見た動く物体を親と認識する。
生後10数時間の経過後以降に、本物の親を見せても、ひなはそれ(本物の親)を親とは認識しない。 つまり、いったん刷り込まれた学習は、修正されにくく、変更されにくい。
このように、発育初期のかぎられた時期にのみ、あることを記憶することを、刷込み(すりこみ、imprinting インプリンティング)という。
オーストラリアのローレンツが、うまれたばかりのガンにガチョウや動く模型などを見せる実験によって、刷り込みを証明した。
なお、ニワトリのひな(ひよこ)にも、刷り込みはある。
知能
編集知能(intelligence)とは、新しい状況に対して、その状況を正しく判断し、適切な行動をとることである。 動物に知能があるかどうか確かめる実験として回り道実験などがある。 これは動物をひもにつなぎ、そのままでは餌に届かないが、回り道をすることで餌に届くようにする。 これにより動物がエサに届く時間で動物の知能を測る方法である。
ヒト以外の動物では、例えばチンパンジーなどが道具を使ったり、単純な言語を扱ったりするなど知能が高いとされている。
情報伝達
編集
[8] 動物の情報伝達には、視覚や聴覚による方法と、フェロモンによる方法などがある。
動物の情報伝達では、ミツバチの8の字ダンスなどが有名である。 ミツバチは、餌の方向・距離を様々なダンスによって他の仲間に伝える。餌が近いときは円形のダンスを、餌が遠いときは8の字のダンスをする。
フェロモン(pheromone)とは、動物の体内で作られ、体外に分泌されて、極微量でも他の個体の行動に影響を与える化学物質のことである。 例えば、アリは餌を見つけたアリは、道しるべフェロモンを出しながら巣へ戻り、他のアリはその道しるべフェロモンをたどり餌にたどり着く。
生物時計
編集生物時計(biological clock)とは、生物の体内の時計機構である。 生物時計は目覚めと睡眠をつかさどり、体温・血圧・心拍数などを調整している。 また、渡り鳥の太陽コンパスも生物時計に基づいている。動物の生物時計は細胞一つ一つにあるが、それらを脳の視交叉上核()が統合している。 アサガオの開花も生物時計が関係している。
脚注
編集- ^ KIM E. BARRETT ほか原著改訂、岡田泰伸 監訳『ギャノング生理学 原著23版 』丸善株式会社、平成23年1月31日 発行、P234
- ^ 吉里勝利ほか『スクエア 最新図説生物』第一学習社、2004年1月10日発行、p.137
- ^ 小澤瀞司・福田康一郎 監修『標準生理学』、医学書院、2015年8月1日 第8版 第2刷、P10、
- ^ 小澤瀞司・福田康一郎 監修『標準生理学』、医学書院、2015年8月1日 第8版 第2刷、P105、
- ^ 吉里勝利ほか『スクエア 最新図説生物』第一学習社、2004年1月10日発行、p.141
- ^ 今井正 ほか『標準薬理学 第7版』 、医学書院、2015年3月25日 第7版 第1刷、P226
- ^ 学習 - Yahoo!百科事典
- ^ 吉里勝利ほか『スクエア 最新図説生物』第一学習社、2004年1月10日発行、p.152
参考文献
編集- 田中隆荘ほか『高等学校生物I』第一学習社、2004年2月10日発行、pp.156-213
- 『NHK高校講座 生物』第22-33回
- 生物学用語辞典 - Weblio 学問
その他、高校の検定教科書などを参照。 [1]
- ^ 吉里勝利ほか『スクエア 最新図説生物』第一学習社、2004年1月10日発行、p.155